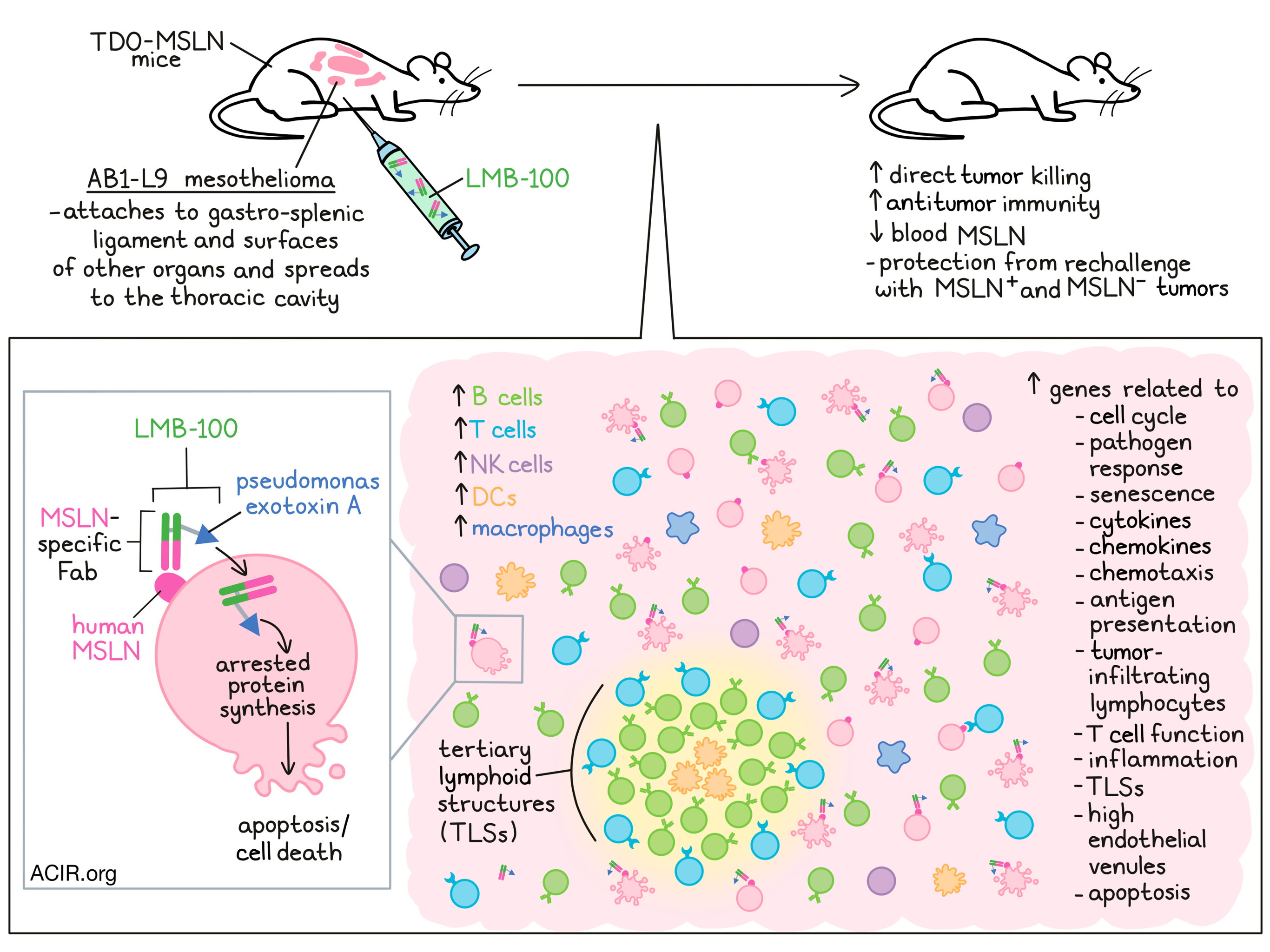
The tumor-associated antigen mesothelin (MSLN) is expressed by mesothelioma and other types of cancer. LMB-100 is an immunotoxin consisting of a Fab recognizing human MSLN that is linked to a toxin (Pseudomonas exotoxin A), which induces direct tumor cell killing by arresting protein synthesis. Liu et al. investigated in a mesothelioma mouse model whether LMB-100 also induces an antitumor immune response, enhancing the therapeutic effects. Their results were recently published in PNAS.
The researchers developed an orthotopic mouse model using TPO-MSLN mice, which express a human MSLN transgene in their thyroid gland, and hence have a normal immune system that does not recognize human MSLN as foreign. Mice were intraperitoneally (i.p.) inoculated with the mouse mesothelioma cell line AB1 expressing hMSLN (AB1-L9). Tumors grew attached to the gastro-splenic ligament and on the surface of other organs, both consistent with peritoneal mesothelioma, and could spread into the thoracic cavity. To assess the treatment activity of LMB-100, one dose was injected i.p. on day 5 after inoculation, which significantly reduced the tumor size and decreased blood MSLN levels.
The researchers then assessed the most effective dosing strategy. First, they compared one dose on day 5 with two doses on days 5 and 9. With one dose, 47% of mice had a CR, while two doses resulted in 50% CR, suggesting the second dose did not have an additional benefit. Testing delayed dosing, survival decreased when the two-dose regimen was given on days 7 and 11 or days 9 and 13. Treatment efficacy was also reduced when one dose was given on day 7 or 9.
The mice with CRs were re-challenged subcutaneously (s.c.) with AB1-L9 cells 1.5 months after initial inoculation to determine if immune memory was formed after treatment. None of the mice that had received one-dose treatment and only 2 of the 19 mice that received the two-dose regimen experienced tumor formation. To assess whether this induced antitumor immunity was directed at MSLN or also at other tumor antigens, the researchers challenged 10 mice with AB1 cells not expressing MSLN, which were rejected in 8 mice, suggesting a response was induced against other tumor antigens.
To study the immune response to therapy, tumor samples were obtained at various time points and subjected to the Nanostring immune profiling. Increases in transcripts at 12 hours were associated with pathways related to the cell cycle, pathogen response, and senescence. These increases peaked 24 hours after treatment and decreased at the 48 hour timepoint. At 24 hours, immune-related pathways were upregulated, including those associated with chemotaxis, antigen presentation, and tumor-infiltrating lymphocytes (TIL). These effects were not caused by anti-drug responses induced by the immunogenicity of LMB-100.
Assessing the most upregulated gene transcripts, five categories were detected related to cytokines and chemokines, inflammation, pathogen responses, apoptosis, and T cell functions. To assess whether this upregulation was induced in tumor cells or other cells in the tumor microenvironment (TME), tumor cells were treated in vitro and analyzed. Of the 98 genes that increased in mice tumors, 40 were increased in tumor tissue, indicating a role of non-malignant cells in the antitumor response.
To assess whether dead tumor cells killed by LMB-100 alone could induce antitumor immunity without the presence of other cells in the TME, Liu et al. treated tumor cells with LMB-100 in vitro before inoculation. As expected, on day 33, all mice inoculated with pre-treated cells were alive and without tumors. However, when these mice were re-challenged s.c. with AB1-L9 cells, none rejected the tumors. These data indicate a critical role for non-tumor cells in the TME for the induction of antitumor immunity.
Assessing changes in tumor-infiltrating immune cells after in vivo treatment, the researchers used RNA analysis and detected a time-dependent increase in B cells, T cells, NK cells, DCs, and macrophages, with the most significant increase in B cells. At 48 hours, there was an absolute increase in NK cells and DCs, CD8+ T and Th1 cells remained unchanged, and there was a large increase in B cells. Histologically, at 48 hours, there was an increase in B cells and CD8+ T cells, and the B cells were found aggregated in tertiary lymphoid structures (TLS) that were distributed throughout the tumor. Regions rich in B cells were surrounded by CD4+ and CD8+ T cells, and regions rich in CD11c+ follicular DC-like cells were found in the B cell-rich core. The number of TLSs increased over time in response to treatment.
Genes associated with TLS, such as Lymphotoxin β, Light, Ccl21a, Ccl19, Ccr7, Cxcl13, and Cxcr5 were upregulated after treatment. These genes were not detected in tumor cells treated in vitro. Furthermore, there was an increase in expression of the lymphangiogenic factor Vegf-c and adhesion molecules Icam-1 and Vcam-1, which are associated with the formation of high endothelial venules, specialized blood vessels found in TLS.
Finally, the researchers used antibody-based depletion to determine the role of TIL in the antitumor responses induced by treatment. CD8+ T cell and B cell depletion shortened survival and decreased the CR rate. CD4+ T cell depletion also shortened survival, but less strongly compared to CD8+ T cell depletion, while depletion of NK cells did not impact outcomes.
These data suggest that treatment with LMB-100 induces direct tumor killing and an antitumor immune response, recruiting TIL to the tumor and inducing TLS formation. Important future work is to establish the critical signal(s) generated by dying tumor cells and their cellular targets. Once proven to show similar mechanisms in patients, these strategies may work synergistically with other immunotherapeutics.
Written by Maartje Wouters, image by Lauren Hitchings



