Migration to and invasion of solid tumor tissue by lymphocytes is essential for antitumor immunity and response to virtually any type of immunotherapy. However, the mechanisms by which lymphocytes enter tumors through vasculature remain largely unclear. Therefore, Asrir, Tardiveau, Coudert, Laffont, and Blanchard et al. studied high endothelial vessels (HEVs) – specialized blood vessels for lymphocyte recruitment in lymphoid organs – in various tumor models. Their results were recently published in Cancer Cell.
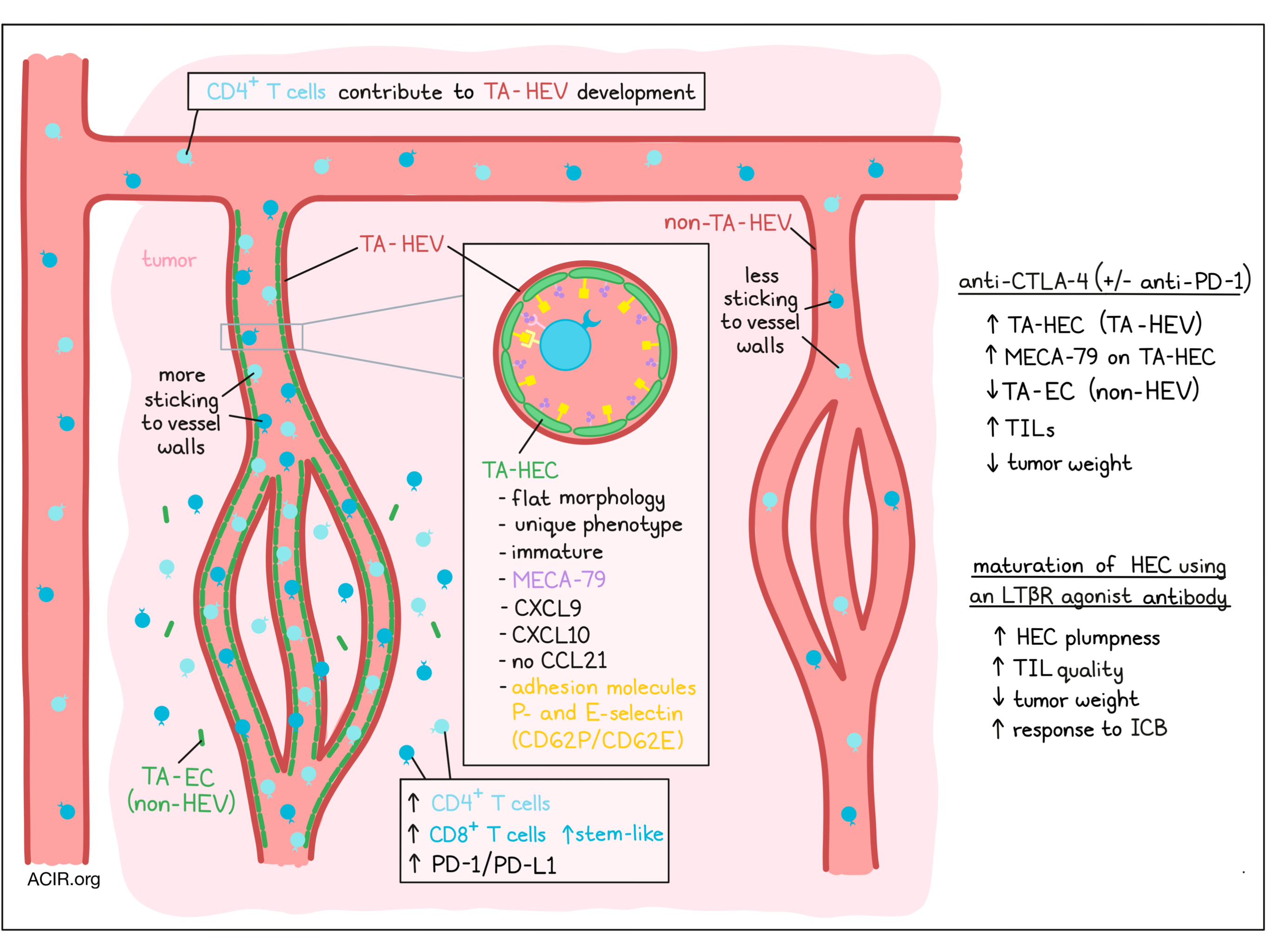
The researchers started by assessing the effects of cancer immunoediting on tumor-associated HEVs (TA-HEVs) using the methylcholanthrene (MCA)-induced cancer mouse model. Immunogenic MCA tumors obtained from Rag2-/- mice regress (MCAreg) when implanted in C57BL/6 WT mice. In contrast, MCA tumors derived from WT mice have undergone immunoediting and form progressive sarcomas (MCAprog) when implanted in WT mice. In both of these tumors, TA-HEVs were detected, and the TA-HEV endothelial cells (TA-HEC) had a flat morphology, in contrast to the plump morphology these cells have in lymph node HEVs. MCAreg tumors had significantly fewer TA-HEC than MCAprog tumors, while they had a higher expression of MECA-79, indicative of a more mature/functional phenotype. TA-HECs were found alongside CD8+ T cell infiltration in both tumors.
RNAseq analysis of various HEC types in healthy and inflamed lymph nodes and TA-HECs revealed a unique TA-HECs phenotype. TA-HECs expressed genes suggesting they derive from post-capillary venule endothelium. These cells had downregulated levels of genes associated with arterial and capillary endothelium and activity-dependent genes, while they had upregulated levels of inflammatory genes. The cells expressed low levels of the lymphotoxin B receptor (LTβR)-dependent genes, which suggests an immature HEV phenotype. TA-HECs expressed genes for CXCL9 and CXCL10, but not CCL21, an important chemokine for T cell recruitment. However, CCL21high cells were found in tumors surrounding TA-HEVs. The HECs also expressed various adhesion molecules (P- and E-selectin) essential for lymphocytes sticking to HEVs.
Adoptive transfer of T cells to MCAprog tumors (in Rag2-/- mice) resulted in an increase of TA-HECs, and depletion experiments showed that CD4+ T cells were essential for the induction of these cells. When mice were treated with anti-CTLA-4 (but not anti-PD-1), the frequency of TA-HECs increased, as well as the influx of (Ki67+ and IFNγ-producing) CD8+ and CD4+ T cells. This increase was associated with the upregulation of PD-L1, and an increased number of PD-1+CD8+ T cells. Treatment with anti-CTLA-4 also caused an increase in both stem-like and terminally exhausted CD8+ T cells.
When anti-CTLA-4 and anti-PD-1 treatments were combined (dual immune checkpoint blockade [ICB]) the strongest effect on tumor growth inhibition was seen, with more mice responding to treatment. Treatment increased the frequency of TA-HECs and reduced the number of non-HEC tumor-associated endothelial cells (TA-ECs). This was associated with higher infiltration of CD4+ and CD8+ T cells, with higher ratios over Tregs, and increased frequencies of stem-like and terminally exhausted CD8+ T cells. Depletion of CD4+ T cells increased the number of TA-ECs and reduced the MECA-79 expression on TA-HECs. This was associated with decreased PD-L1 expression and CD45+ cell counts, and increased tumor weight. Furthermore, when mice were treated with the S1P receptor inhibitor FTY720 to limit lymphocyte recirculation, the influx of lymphocytes, increase in PD-L1 expression, and decrease in TA-ECs induced by combination checkpoint blockade was averted.
The researchers then performed in vivo imaging to assess the role of TA-HEVs in lymphocyte infiltration into MCAprog tumors. When mice were treated with combination ICB, the percentage of lymphocytes found rolling along the walls of intratumoral blood vessels was 40% in the TA-HEVs and less than 10% in non-HEV blood vessels. Blockade of the lymphocytes and endothelial receptors using anti-CD62L (aka L-selectin) and anti-PSGL-1 (the ligand for P-selectin) antibodies showed that these pathways were essential for the tethering and rolling interactions of lymphocytes with TA-HEVs. The sticking fraction – the number of cells that went from rolling to firm arrest in the vessels – was almost entirely restricted to TA-HEVs.
To assess the next steps in the lymphocyte extravasation cascade, the researchers used multiphoton in vivo imaging. They noticed that after the arrest, lymphocytes crawled on the endothelium before entering the tumor tissue. ICB treatment increased the lymphocyte homing, and when the CD62L and PSGL-1 antibodies were used, it prevented the influx of T cells in response to ICB.
Given that TA-HEVs are such an important site for lymphocyte extravasation, and the finding that TA-HECs had an immature phenotype, the researchers assessed the effect of maturation of TA-HECs. Treatment with an LTβR agonistic antibody alone resulted in an increase in the number and the maturation of TA-HEC cells (increased MEC-79A surface expression). This was associated with reduced tumor weight, but did not affect overall CD8+ T cell infiltration. However, there was a decrease in the number and frequencies of exhausted and terminally exhausted CD8+ T cells, and an increase in the proportion of stem-like CD8+ T cells. When the LTβR agonistic antibody treatment was combined with the dual checkpoint blockade, there was also enhanced maturation of TA-HECs, which was associated with increased numbers of stem-like CD8+ T cells and PD-L1 expression, though it did not affect the number of terminally exhausted CD8+ T cells. The more mature TA-HEVs were found in the periphery of tumors, and the endothelial lining had a plump morphology, like the cells in lymph node HEVs. More mice responded to the triple treatment than to ICB treatment alone.
Finally, the researchers assessed whether TA-HEVs may also be predictive for therapy response in a cohort of 93 patients with unresectable stage III or IV metastatic melanoma treated with either anti-PD-1 alone or in combination with anti-CTLA-4. Pre-treatment tumors had TA-HEVs, which were associated with CD8+ T cell infiltration. Responders to the combination checkpoint blockade treatment had higher TA-HEV expression scores; however, this was not the casen those treated with anti-PD-1 alone. In dual ICB-treated patients, those with a high TA-HEV score had a better response to treatment, resulting in improved overall and progression-free survival.
These data suggest the importance of TA-HEVs for TIL infiltration and response to checkpoint blockade involving anti-CTLA-4. Additionally, these data highlight the translational potential to further improve ICB therapy efficacy by enhancing the number and/or maturation of tumor-associated high endothelial cells.
Write-up by Maartje Wouters, image by Lauren Hitchings
Meet the researcher
This week, lead author Jean-Philippe Girard, answered our questions, on behalf of the five co-first authors: Assia Asrir, Claire Tardiveau, Juliette Coudert, Robin Laffont and Lucas Blanchard.

What prompted you to tackle this research question?
I have been studying high endothelial venules (HEVs), specialized blood vessels for lymphocyte entry in lymph nodes, since my post-doc with Timothy Springer at Harvard Medical School in the ‘90s. When I came back to France, I decided to define the mechanisms regulating HEV formation, in order to be able, one day, to induce HEVs in tumors. In 2011, I had the great surprise to discover that blood vessels with HEV characteristics (now called tumor-associated HEVs or TA-HEVs) are frequently found in human solid tumors (melanomas, breast, ovarian, colon, and lung carcinomas). Unlike classical tumor blood vessels, TA-HEVs correlated with T and B cell infiltration and favorable clinical parameters. These observations suggested that TA-HEVs mediate lymphocyte infiltration into tumors. In the current work, we visualized for the first time lymphocyte extravasation through TA-HEVs, and we demonstrated that TA-HEVs are the major sites of lymphocyte entry into tumors during cancer immunotherapy.
What was the most surprising finding of this study for you?
First, we didn’t expect that TA-HEVs would be the almost exclusive sites of lymphocyte entry into tumors during immune checkpoint blockade. We believe that these observations are explained by the unique bifunctional hybrid phenotype of endothelial cells lining TA-HEVs. Second, we were surprised that TA-HEVs were highly predictive for better response and survival of metastatic melanoma patients treated with anti-PD-1/anti-CTLA-4 combination, but not those receiving anti-PD-1 only. However, these findings are consistent with the distinct mechanisms of anti-PD-1 and anti-CTLA-4 antibodies. James Allison, Padmanee Sharma, Antoni Ribas, and others have shown that anti-CTLA-4 treatment increases T cell infiltration into tumors. Anti-CTLA-4 efficacy may thus be more dependent on the presence of TA-HEVs than anti-PD-1 therapy, which is believed to act primarily at the tumor site. Nevertheless, TA-HEVs are likely to be also important for anti-PD-1 treatment because they mediate lymphocyte entry at baseline, and lymphocyte infiltration is a prerequisite for anti-PD-1 efficacy. Moreover, there is growing evidence that anti-PD-1 also acts in the periphery.
What was the coolest thing you’ve learned (about) recently outside of the lab?
Recently, I had the chance to swim with manta rays. I am totally fascinated by mantas. They are so graceful and peaceful. Their encounter is always a magical moment. One of the mantas passed very close to me. It was a fantastic experience. Such encounter illustrates perfectly the inquisitive nature of these charismatic creatures. I learned that the latest genetic analyses indicate that there are at least two distinct species of manta rays around the world. Sadly, mantas are increasingly threatened by human activities. Global conservation efforts are needed to save the mantas and the other magical creatures of the oceans.



