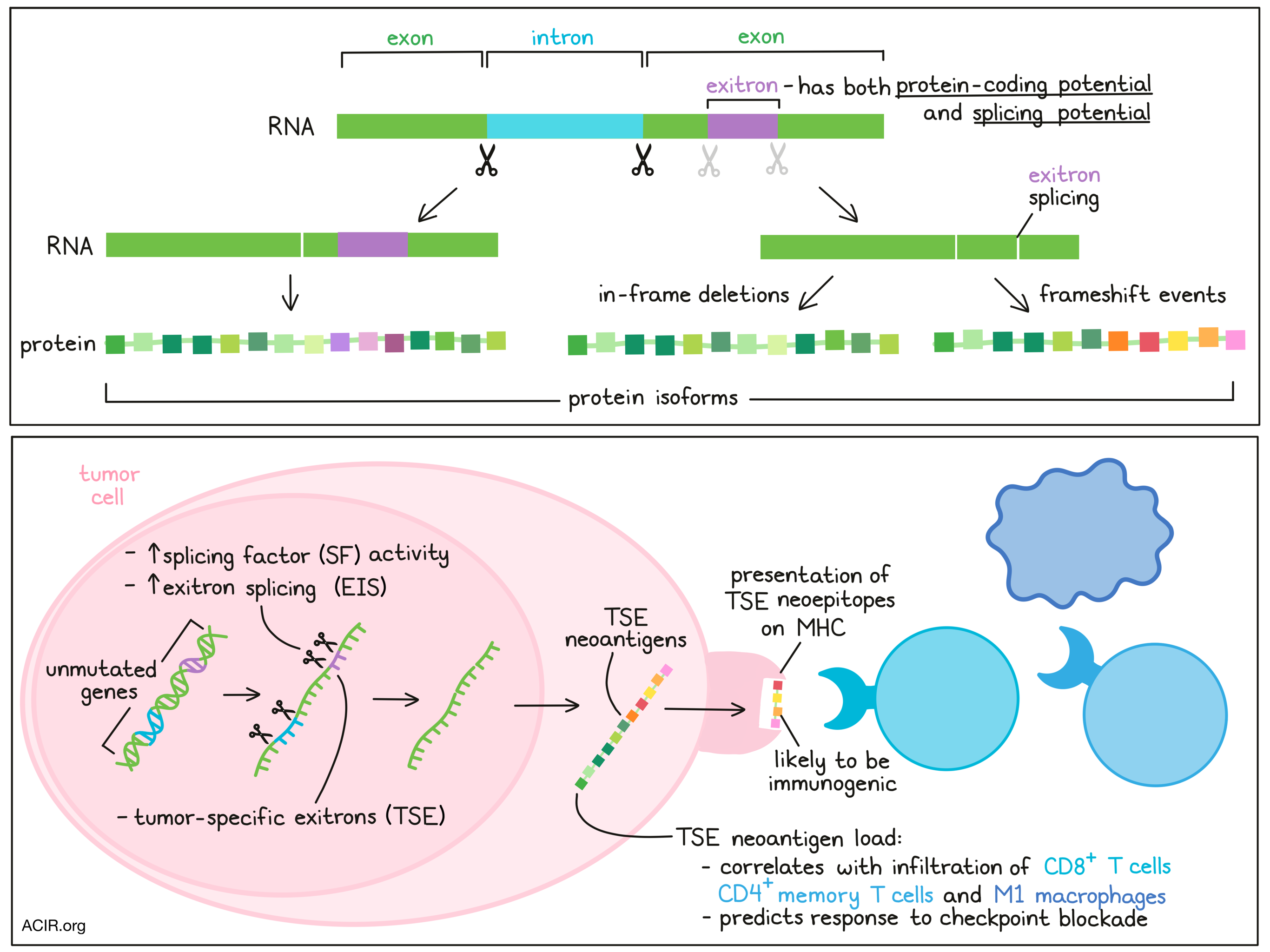
Exitrons are internal regions of an exon that have both protein-coding and splicing potential. When these exitrons are spliced, it may lead to protein isoforms, including in-frame deletions, as well as out-of-frame neo open reading frames. As not much is known about the abundance or impact of exitrons in cancer, Wang and Liu et al. quantitatively assessed exitrons in cancer using bioinformatics analyses, revealing functional roles, as well as a new potential source of neoantigens. Their results were recently published in Molecular Cell.
The researchers started by assessing RNAseq data from 9,599 patients with 33 types of cancer from TCGA using their bioinformatic tool ScanExitron. The tool identified 129,406 exitrons in 39,755 exons, accounting for 14.8% of the GENCODE human exome. Exitrons had distinct cis-acting features, including weak 5’ and 3’ splice sites, high GC content, and short length, and were mainly found in medium-sized (100 bp - 1kbp) exons. In healthy tissue obtained from the GTEx RNAseq dataset, exitrons were spliced in 17.1% of genes – much lower than in tumor tissue, where they were spliced in 62.7% of genes.
The highest numbers of exitron splicing (EIS) events were found in ovarian, esophageal, and stomach cancer, and acute myeloid leukemia (AML). To compare the EIS burden between tumor and matched healthy tissues, eight tumor types were selected with at least 40 tumor and healthy tissue pairs. There was a significantly higher EIS burden in the tumor samples, and there were 16 EIS events that were spliced differently between matched tumor and healthy tissue. These exitrons impacted genes implicated in cancer, such as loss-of-function of the tumor suppressor genes FOXO4 and SPEN.
Exitron splicing has been associated with dysregulation of splicing factor (SFs), and by clustering cancer types based on the median expression of 404 SF genes, the cancer types with a high EIS burden clustered together. Additionally, within cancer types, the activity of SFs correlated with EIS burden. Using a generalized additive model, the researchers found that EIS dysregulation in cancer tissue could largely be explained by differential expression of SFs. Functionally implicated SFs were genes coding for the spliceosome and proteins regulating RNA transport and mRNA surveillance.
The researchers then identified tumor-specific exitrons (TSEs), defined as exitrons found in tumors that were not spliced in the healthy GTEx samples and were spliced in 3 or fewer normal TCGA samples. Ovarian, esophageal, and stomach cancer, and AML had the highest number of TSE-splicing events. To assess the clinical impact of TSEs, those TSEs that were spliced in ≥10 samples and were associated with survival were defined as clinically informative exitrons. These clinically informative exitrons were found in 21 cancer types out of 25 assessed in TCGA.
Given that some TSE-splicing events were found in multiple cancer types, Wang and Liu et al. investigated their functional role by assessing the enriched TSE-splicing events in each cancer type. Ranking these significantly exitron-spliced genes (SEGs) based on their frequency showed that genes in the COSMIC cancer gene census were significantly enriched. Some of the highest-ranked SEGs were members of the FET protein family, which is involved in transcriptional regulation and RNA processing. Patients with these TSEs had shorter progression-free survival, suggesting these genes may play a role in cancer progression.
To determine whether any of the SEGs could regulate cancer driver genes, a tissue-specific analysis was performed in prostate cancer. One of the SEGs found was NEFH, which encodes heavy neurofilament protein. Expression of this gene was lower in tumors than in benign samples in TCGA, and inversely correlated with the Gleason score, a marker for prostate cancer aggressiveness. Given that NEFH rarely has mutations in prostate cancers, the researchers then investigated whether SEGs generally lack mutations. Indeed, significantly mutated genes (SMGs) and SEGs bifurcated in a volcano plot analysis. This mutual exclusivity was confirmed when genes with somatic mutations were compared with TSE splicing, and was also seen at the patient level. Mutation burden and EIS burden were independent across all cancer types.
Given that EIS affects protein-coding exons, there is a chance that these lead to frameshift events resulting in neoantigens. Using ScanNeo and netMHCpan on the TCGA data, the researchers identified 1.4 neoantigens per TSE-splicing event, with the highest burden of TSE neoantigens found in ovarian cancer. Using MS-GF+, a search tool for proteomics datasets, to search mass spectrometry data in 32 ovarian and 35 breast-invasive carcinoma samples from the Clinical Proteomic Tumor Analysis Consortium project for these predicted epitopes, the researchers identified 28 neoepitopes that were derived from TSE splicing. To determine whether these neoepitopes could be processed and presented by MHC, a separate cohort of 11 ovarian samples with data on transcriptome profiling and immunoaffinity purification of MHC complexes followed by MS analysis was examined. This confirmed that 26 TSE neoantigens could be presented by MHC. These TSE neoantigens had a high potential to be recognized by CD8+ T cells, based on a high hydrophobic content, as has been reported for other neoantigens, and had longer neo open reading frames (neoORFs), suggesting high immunogenicity.
In the TCGA data, a positive correlation was found between TSE neoantigen load and higher infiltration of CD8+ T cells, M1 macrophages, and CD4+ memory T cells. To assess whether TSE neoantigen load was also associated with checkpoint blockade response, the data from three cohorts of patients with melanoma and one cohort of patients with clear cell renal cell carcinomas (ccRCC) who received CTLA-4 or PD-1 checkpoint blockade were assessed. A correlation between TSE and TSE neoantigen burden and response was found only in ccRCC, and TSE neoantigen load was a better predictor of outcome than other response signatures, such as mutation neoantigen load, CD8+ T cell infiltration, and PD-L1. TSE neoantigen load was also highly correlated with expression of T cell, cytolytic activity, and immunoregulatory markers in ovarian and clear cell renal cell carcinoma.
The findings in this study suggest that exitrons in cancer cells provide multiple novel perspectives, including contributions to the oncogenic phenotype, identification of new cancer driver genes, and sources for a new set of neoantigens that are potentially targetable with immunotherapy. It will be of particular interest to determine whether a high TSE neoantigen burden can stratify patients with ovarian cancer who might benefit from checkpoint inhibition, which has generally been of limited success in this cancer type.
Write-up by Maartje Wouters, image by Lauren Hitchings
Meet the researcher
This week, lead author Rendong Yang answered our questions.

What prompted you to tackle this research question?
We coincidentally discovered frequent exitron splicing events in prostate cancer patients when examining expressed genetic insertions and deletions as reported in our previously published study in BMC Genomics 2018. We realized exitron splicing events potentially change the protein sequences in a similar way as genetic mutations. Because genetic mutation is a known source of cancer driver genes and neoantigens, this led us to think about if exitron splicing events are prevalent in cancer transcriptomes, and whether they could be leveraged for cancer driver events and neoantigen discovery.
What was the most surprising finding of this study for you?
We discovered that exitron splicing are mutually exclusive with somatic mutations. This led us to discover previously undetected cancer driver genes and exitron splicing-derived neoantigens. Mostly importantly, most of the detected exitron splicing events are tumor-specific, and neoantigens derived from these events could be highly immunogenic compared to those derived from mutations. In addition, several studies in different tumor types showed a correlation between high tumor mutation burden (TMB) and response to immune checkpoint blockade (ICB) therapy, likely due to the increase in mutation-derived neoepitopes. Intriguingly, we found that the correlation between exitron splicing-derived neoepitope load and ICB efficacy was more pronounced in tumors with low TMB, such as clear cell renal cell carcinoma. This suggests RNA splicing-derived neoantigens are potential predictive biomarkers for ICB response in tumors with low TMB.
What was the coolest thing you’ve learned (about) recently outside of work?
My most enjoyable time is when I do garden work together with my family. We have planted a few different kinds of vegetables in our garden, such as cucumber, green beans, and tomatoes. This is the third year we have been planting these vegetables, and this year they all turned out to be so good. We planted the seeds almost at the same time, but the seeds have their own life cycles. When I saw the green beans and tomatoes are all having sprouts coming out, but the cucumbers were still so quiet and had nothing show up, I almost gave up and dug it out to see what happened to them. Fortunately, my wife said we should wait one more week to see if it works. Finally, the cucumbers are all coming out and are growing well now. Every seed has its own life cycle. Beyond water, soil and sunshine, the most important factor I learned is being patient.



