Blocking the PD-1/PD-L1 axis using anti-PD-L1 antibodies is a commonly employed immunotherapeutic strategy to prevent PD-1-mediated inhibition of T cells by tumor cells. Suspecting that the mechanism of action of PD-L1 blockade might be more complex, however, Mayoux et al. investigated the role of PD-L1-expressing dendritic cells (DCs) in PD-L1 blockade. The results were recently published in Science Translational Medicine.
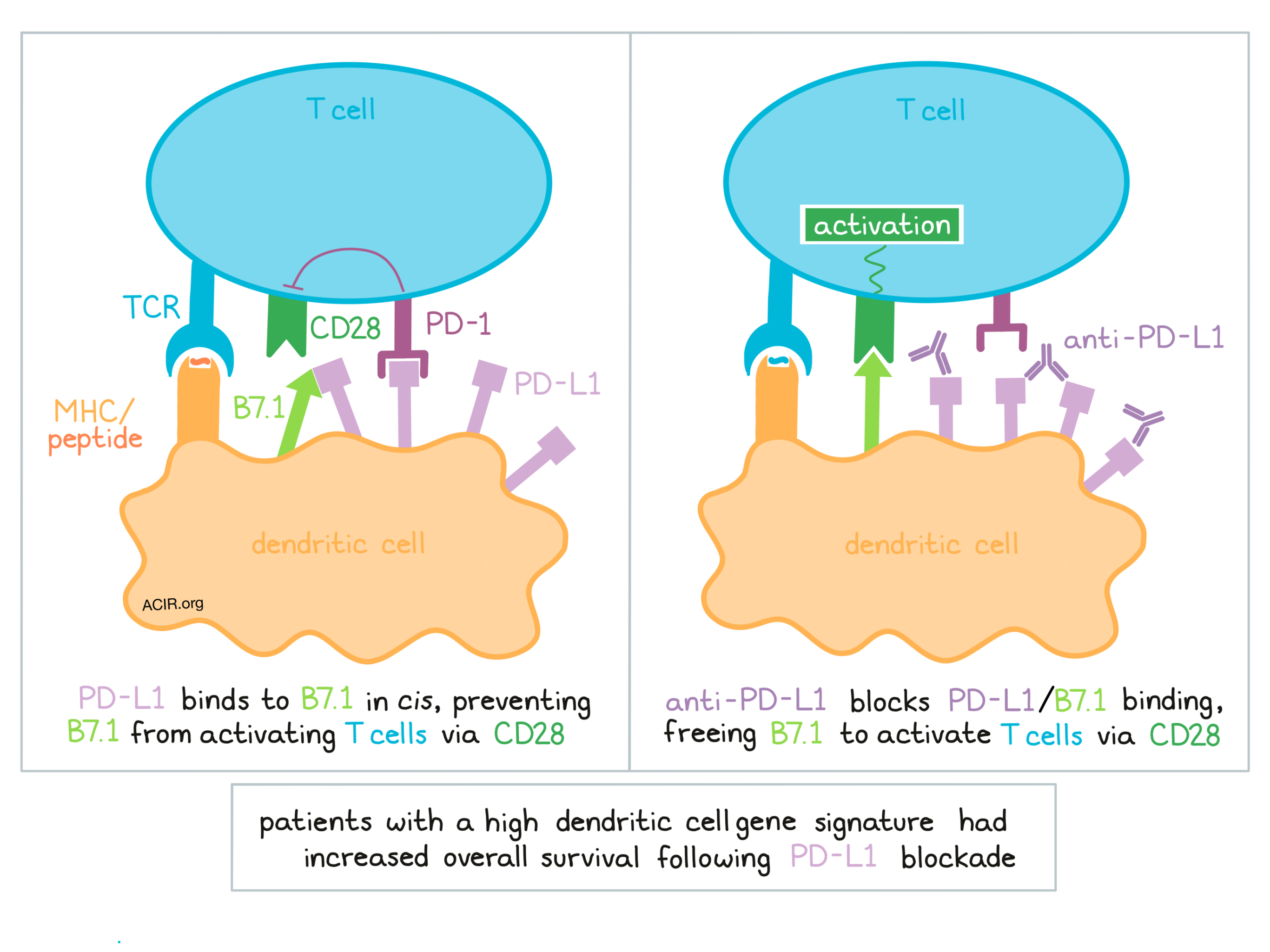
When a DC primes a T cell, the generally understood mechanism is that B7.1 on dendritic cells engages with the positive costimulator CD28 on T cells, encouraging T cell activation. If the negative costimulator PD-1 is also engaged on T cells, then T cell activation could be inhibited. Mayoux et al. investigated the expression of PD-L1, PD-1, and the CD28 stimulatory ligand B7.1 (aka CD80) on various DC subsets from patients with lung cancer and found that both tumor-associated and peripheral DCs expressed PD-L1. They also observed that tumor-associated but not peripheral cross-presenting DCs (CD141+ and CD1c+) expressed B7.1. Focusing on a double-positive (PD-L1+ B7.1+) population in cross-presenting DCs from fresh renal cell carcinoma samples, and cognizant that PD-L1 and B7.1 have been known to interact in cis, the researchers used an antibody binding assay to quantify surface expression and observed that PD-L1 was 20-fold more abundant than B7.1 on both CD141+ and CD1c+ conventional dendritic cells.
Based on this evidence, Mayoux et al. hypothesized that the abundant PD-L1 could sequester the less abundant B7.1 in cis on DCs, leaving excess PD-L1 available for binding. To test this, they used monocyte-derived DCs from healthy donors in an in vitro system to develop LPS-matured DCs and observed expression patterns of PD-L1 and B7.1 that were similar to those observed in tumors. In a confocal imaging analysis, when the DCs were observed forming synapses with T cells, PD-1 and CD28 colocalized more often than PD-1 and PD-L1, consistent with recent findings that PD-1 signaling may be involved in the dephosphorylation of CD28 at the immunological synapse. Interestingly, B7.1 was barely observed colocalizing with CD28 at all, instead interacting to a much higher degree with PD-L1 in cis, likely due to its higher binding affinity.
To test whether blocking PD-L1 might release B7.1 from sequestration and make it available for interaction with CD28, the researchers modeled the physical interactions between B7.1/PD-L1 and B7.1/CD28. First, they showed that anti-PD-L1 binds to PD-L1-expressing cells with a binding affinity 700-fold higher than the binding affinity between PD-L1 and B7.1, suggesting that the antibody would readily disrupt PD-L1/B7-1 binding. Next, using cells engineered to mimic the expression of B7.1 and PD-L1 on DCs, and CD28-Fc to mimic T cells, the researchers observed an increase in B7.1/CD28 interactions when anti-PD-L1 was present. These results were confirmed in a primary cell-cell system, which also showed that pre-incubating mature DCs with anti-PD-L1 enabled stronger interactions between B7.1 on DCs and CD28 on T cells. Together this evidence indicated that anti-PD-L1 can disrupt the interatctions between PD-L1 and B7.1 in cis, freeing up B7.1, enhancing B7.1 binding to CD28, and activating T cells.
To measure the functional consequences of anti-PD-L1 disrupting the binding between PD-L1 and B7.1 in cis on DCs, Mayoux et al. evaluated CD28 downstream signaling in Jurkat reporter cells. To better isolate the specific contribution of blocking the interaction between PD-L1 and B7.1 in cis, Jurkat cells were first incubated with anti-PD-1 to block PD-1/PD-L1 binding in trans, which increased CD28 signaling by 32%. The cells were then incubated with anti-PD-L1, which further enhanced CD28 signaling by 20%, for an overall increase of 52% in CD28 binding. When the researchers repeated the experiment using a PD-L1-targeting antibody that does not substantially block PD-L1/B7.1 interactions, the increase in CD28 signal transduction was significantly lower.
Next, Mayoux et al. studied the consequences of pretreating activated human DCs with PD-L1 blockade and found that it directly enhanced their ability to stimulate the proliferation of allogeneic T cells. To study whether this effect could generate de novo immune responses, mouse splenic CD11c+ DCs were cocultured with DQ-OVA (cross-presented as the SIINFEKL peptide) and treated with anti-PD-L1. In coculture with naive T cells from OT-I mice, the DCs pretreated with anti-PD-L1 induced more T cell proliferation; increased T-bet and Eomes in T cells, suggesting enhanced T cell effector potential; and enhanced production of granzyme B in T cells in response to target cells. Similar results observed using SIINFEKL peptide-pulsed DCs pretreated with anti-PD-L1.
To test the extent to which tumor-associated DCs contribute to responses to PD-L1 blockade in a clinical setting, Mayoux et al. established a DC gene signature based on RNAseq data and quantification of expression of key genes that defined DC subsets. In patients with renal cell carcinoma treated with atezolizumab (anti-PD-L1), patients with a high pretreatment DC signature showed better overall survival. In patients with non-small cell lung cancer treated with either anti-PD-L1 or docetaxel, a high DC signature correlated with increased overall survival only in patients in the anti-PD-L1 treatment group. Further, the survival benefit was only observed when PD-L1 was expressed on immune cells, refining the correlation between a high DC signature and increased overall survival, and suggesting that these factors could be considered when making decisions regarding treatment options.
Overall, Mayoux et al. show that the activation of T cells by antigen-presenting cells involves a complex network of interactions between PD-1, PD-L1, B7.1, CD28, and possibly CTLA-4 (not yet investigated). They also show that intratumoral DCs expressing PD-L1 and B7.1 likely play a significant role in the outcome of PD-L1 blockade. In addition to blocking interactions between PD-L1 and PD-1, anti-PD-L1 antibodies also prevent interactions between PD-L1 and B7.1 in cis on dendritic cells, freeing up B7.1 to interact more readily with CD28 on T cells and prime T cell activation and antitumor responses – a mechanism not available to anti-PD-1 antibodies. Importantly, the presence of a DC signature could help guide treatment decisions in the clinical setting.
by Lauren Hitchings



