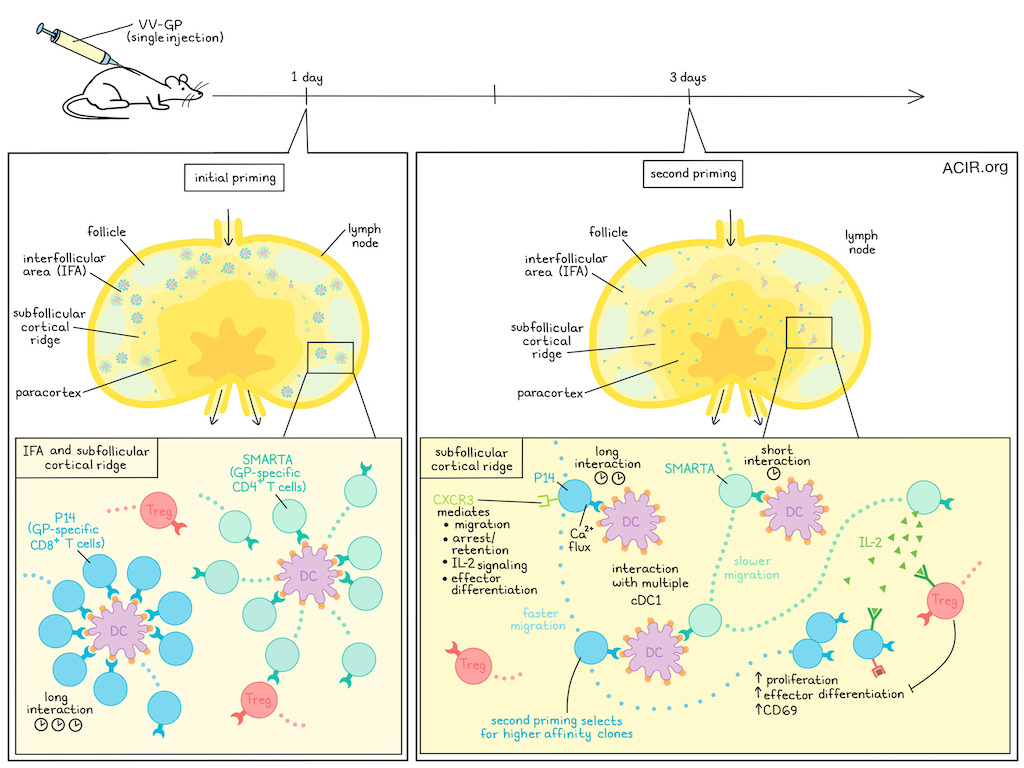
After initial priming through interaction with dendritic cells (DCs), CD8+ T cells disengage and migrate, but little is known about how cytokine signals are transmitted or received by these cells for differentiation during this migration. Jobin, Seetharama, et al. visualized the dynamics of antigen-specific CD8+ T cells, as well as CD4+ T helper (Th) and regulatory (Treg) cells after initial activation. Their novel findings on a second priming phase were recently published in Science.
The researchers began by assessing in which microenvironments viral antigens are presented by DCs in draining lymph nodes (dLNs) after viral infection. Mice were infected with vaccinia virus (VV) expressing a lymphocytic choriomeningitis virus glycoprotein (VV-GP), and GP-specific TCR transgenic CD8+ T cells (P14) were transferred. Within 24 hours of infection, P14 T cells clustered in the interfollicular area (IFA) and cortical ridge areas of the dLNs, where initial priming took place during long-term interaction with DCs. Three days after infection, P14 cells had proliferated and were distributed throughout the paracortex of the dLNs. Further, cells were actively migrating in the interfollicular area.
To further assess the dynamics, naive P14 cells were transferred at the 3-day time point in this viral model. Eight hours post-transfer, T cells had upregulated CD69 and formed clusters. Antigen-presenting DCs were absent from the IFA, but present in the subfollicular cortical ridge area (T cell zone), where antigen presentation was taking place at day 3 post-infection.
Intravital imaging showed hours-long interactions between the P14 cells and cDC1s in the subfollicular areas. At day 3, large P14 cells (suggestive of lymphoblasts) stopped migrating and interacted with cDC1s for at least an hour. The researchers hypothesized that these secondary tight cellular interactions between T cell blasts and cDC1s in the subfollicular LN area shaped the CD8+ T cell response, and named the process the second priming phase. The T cell blasts interacted with multiple local cDC1s during this phase, while in the initial priming there was a clustering of T cells around one DC. Further, the T cells that re-engaged with DCs proliferated during this period.
To investigate the role of CD4+ T cells during the second priming phase, GP-specific SMARTA CD4+ T cells were co-transferred with P14 CD8+ T cells in a similar experimental setting. SMARTA lymphoblasts were also found in the P14 cell niches, but they showed a different migratory pattern. SMARTA cells stopped migrating and interacted with DCs for approximately 10 minutes, after which they migrated at low speed, moving between DCs. Tregs, on the other hand, did not stop migrating and continued to move at similar speeds as during homeostasis.
Productive immunological synapse formation during antigen-specific cell interactions requires Ca2+ mobilization. In vitro experiments with in vivo-activated P14 cells demonstrated that some cells retained their ability to mobilize Ca2+, indicating that they could respond to TCR stimulation, which is required for cellular arrest. In the mouse model, arrested P14 cells in the subfollicular niches showed Ca2+ flux, suggestive of retention of the capability to respond to TCR signaling at day 3 post-infection.
The researchers then determined which chemokine receptors were involved in the recruitment of T cells to DCs and allowed their long-term interactions. In a model infected with VV-OVA, WT and CXCR3 knockout (KO) OT-1 cells were compared. WT cells arrested migration more commonly and longer than the KO cells, suggesting CXCR3 was important for the migration and retention of CD8+ T cells in the subfollicular hubs during the second priming phase. Further, the KO cells did not differentiate into effector T cells (Teff), while the WT cells did. On day 3, the KO cells had reduced levels of pSTAT5 compared to WT cells, suggesting limited IL-2 signaling.
Since Tregs control immune responses by limiting IL-2 availability, the role of Tregs in this process was further assessed. Treg depletion increased the number of virus-specific OT-1 cells expressing the effector marker KLRG1 on day 8 after VV-OVA infection. Further, OT-1 cells expressed increased levels of pSTAT5 on day 3 postinfection. Further experiments showed that Treg-mediated control of these virus-specific T cell responses did not require CXCR3 expression on the Tregs. Tregs had increased levels of gene expression related to activation of IFN- and IL-2-dependent gene pathways, and the elevated levels of IL-2 signaling signals were primarily observed in “resting” Tregs, a subpopulation found during homeostasis in control lymph nodes with low IL-2 signaling signature. In the absence of Tregs, Teff cell generation from CXCR3 KO OT-1 cells was restored. Therefore, Tregs may limit CD8 effector differentiation during the second priming phase.
The researchers then determined the cellular source of IL-2 during the second priming phase using various models. In mice in which CD8+ T cells were unable to produce IL-2, no difference in effector differentiation was observed after infection. When CD4+ T cells were depleted, OT-1 cells had lower levels of pSTAT5 on day 3 post-infection. IL-2Rα-deficient OT-1 cells had reduced expansion and effector differentiation. In a newly developed model in which only CD4+ Th cells were depleted, but not Tregs, an over 5-fold reduction in the number of virus-specific CD8+ T cells was observed on day 8 after infection. Therefore, CD4+ Th cells may play an important role through production of IL-2 in establishing the quantity of antiviral CD8+ T cell responses.
Finally, the researchers assessed whether the second priming phase provided a mechanism for selecting high-affinity clones for further expansion. Comparing the dynamic behavior of low-affinity OVA-specific CD8+ T cells (OT-3) with their high-affinity counterparts (OT-1) showed limited differences during the initial priming phase in terms of migratory arrest, CD69 expression, and proliferation. However, during the second priming phase, the low-affinity OT-3 cells did not engage long-term with DCs and expressed limited pSTAT5. This resulted in 10-fold fewer OT-3 cells and 100-fold fewer Teff cells on day 8 postinfection.
Together, the data suggest the presence of a second priming phase in which CD8+ T cells re-engage with DCs long-term to get signals for effector differentiation. This process relies on signals from CD4+ Th cells and is regulated by Tregs. These data may provide clues for research into improving the efficacy of vaccines and cellular therapies.
Write-up by Maartje Wouters, image by Lauren Hitchings
Meet the researcher
This week, co-first authors Kasia Jobin and Deeksha Seetharama answered our questions.

What was the most surprising finding of this study for you?
We were both very surprised when we noticed that CD8+ T cells re-clustered for a second time after their initial activation. Even more, to see that antigen-specific CD4+ T cells showed a different migration pattern, which again differed from regulatory T cells. Although we had concrete hypotheses about what was happening at this time after the infection, we had to throw them all overboard. It is a great feeling when you observe an unknown process for the first time. Nevertheless, it took us a long time to put all the pieces of the puzzle together, because our results were contradicting paradigms that had long been taken for granted.
What is the outlook?
In principle, we show that the activation of CD8+ T cells is a cyclical process that repeats itself every 2-3 days. After an acute infection, at least in our model, this involves two phases, as the duration of antigen presentation is limited. However, we hypothesize that the principles of cellular interactions which we have discovered for an acute infection also apply to chronic infections and cancer, in which antigen-presentation is continuous. This would have direct implications on treatment regimens for checkpoint immunotherapy or for CAR T cell therapy. We are currently working on precisely these questions. Overall, we are seeing a certain renaissance in research into the role of CD4+ helper T cells in tumor defense, which is only partly based on their cooperation with cytotoxic CD8+ T cells. We believe that the next few years will bring many exciting new findings and therapeutic approaches in this field of research.
What was the coolest thing you’ve learned (about) recently outside of work?
KJ: I’ve been living in Germany for some time and I’m still learning the language. At my home we speak English and Polish. A few nights ago, I was putting my 3-year-old to sleep, and he said: Ohrläppchen. I said: what´s that? He touched my ear lobe and repeated: Ohrläppchen. So, I’ve learned a cool word from a cool little guy.
DS: Recently I travelled to South Korea and learned to properly use chopsticks and make traditional Kimchi. It was exciting to learn their culture and have a well earned break after the paper.



