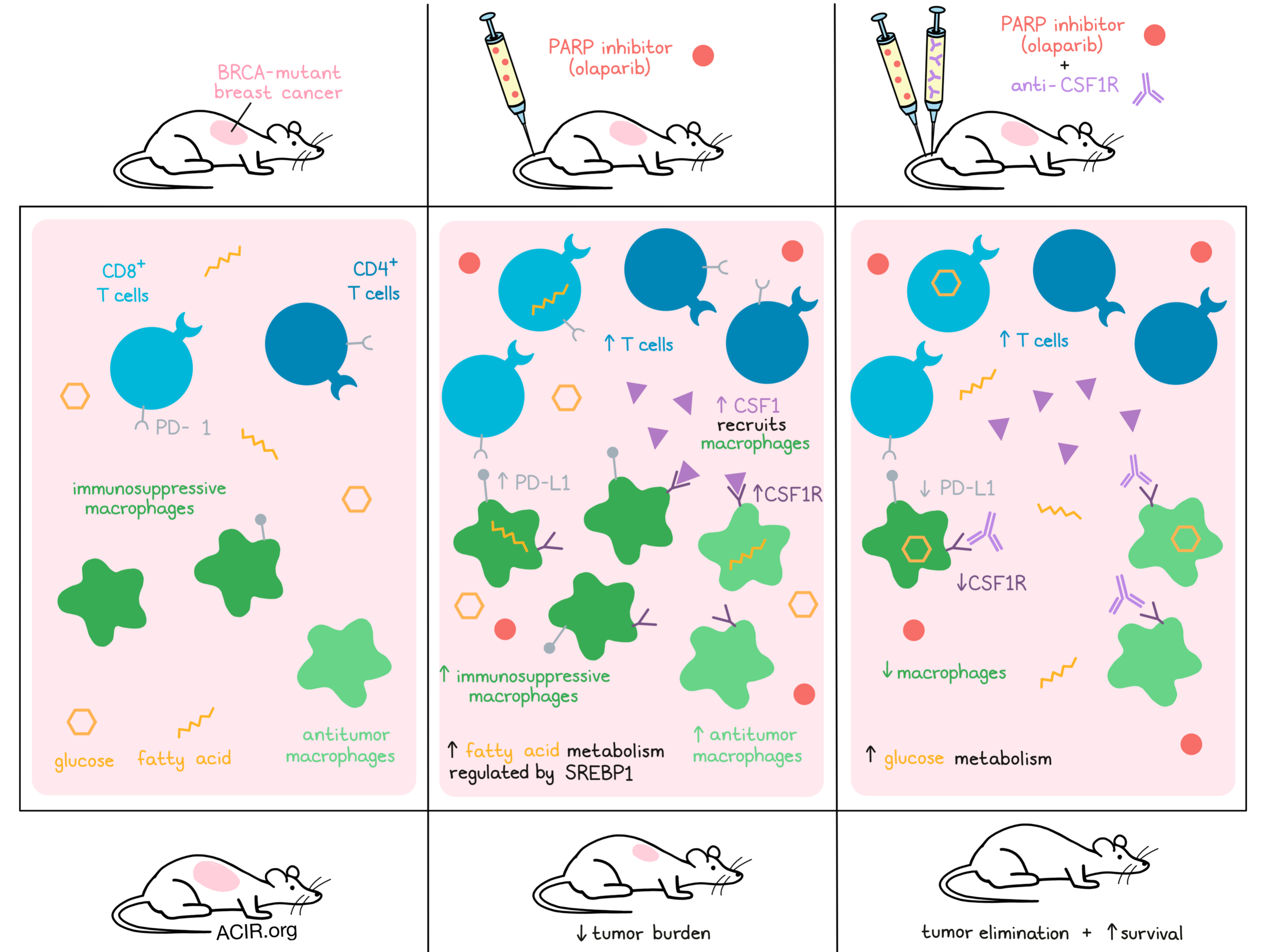
Even though FDA-approved poly(ADP-ribose) polymerase inhibition (PARPi) can be very effective in BRCA-mutant breast cancer, responses are not durable, prompting a need for combination therapies. To provide a mechanistic basis for considering immunotherapeutic strategies, Mehta et al. used high-dimensional single-cell and multi-omics profiling in patient samples and in mouse models to study changes in macrophage and T cell populations following PARPi therapy, and tested approaches to manipulate the macrophages. Their encouraging results were recently published in Nature Cancer.
The researchers started by comparing the immune infiltrate phenotypes of BRCA1-associated and BRCA WT TNBC in tissue sections using multiplex imaging. The BRCA1-mutant tumors were infiltrated with more granzyme B+PD-1+CD8+ and PD-1+CD4+ T cells (including Tregs) than BRCA WT tumors. The most abundant tumor-infiltrating were macrophages, and the BRCA1-mutant tumors had more CD68+ and CD163+ cells, expressing higher levels of PD-L1, indicative of an immunosuppressive phenotype.
To study the effects of the PARP inhibitor olaparib, the K14-Cre;Brca1f/f;p53f/f murine model of TNBC was used. In these mice, olaparib slightly reduced tumor burden and increased CD45+ cell infiltration. Like in the human samples, most infiltrating CD45+ cells were CD11b+F4/80+ mature macrophages, and treatment also induced small increases in T cell infiltrates.
Using Nanostring gene expression profiling in the mouse model, increases in macrophage and myeloid cell-related markers such as Itgam (CD11b), Cd68, Csf1r, and the co-stimulatory molecules Cd80 and Cd86, as well as Cd274 (PD-L1) and Itgax (CD11c) were found. Additionally, genes involved in antigen presentation, chemokine and cytokine signaling cascades, and TLRs, were upregulated, suggesting increased myeloid function. Csf1 levels increased as well with treatment, consistent with previous reports that radiation and chemotherapy increase CSF1 levels, resulting in the recruitment of macrophages.
The researchers then assessed the phenotype of F4/80+CD45+ macrophages after PARPi. The co-stimulatory molecules CD80 and CD40 increased, suggesting induction of an antitumor phenotype, while levels of CD206 (associated with a pro-tumor phenotype) did not change. There was also an induction of the STING pathway effector TANK-binding kinase-1 (TBK1) in macrophages. On the other hand, there were more F4/80+CSF1R+PD-L1+ macrophages and these cells had a higher expression of PD-L1, suggesting overall that both pro- and anti-tumor macrophages increased after PARPi.
To confirm the mouse data in human cells, human monocytes from healthy donors were differentiated into macrophages ex vivo, while being treated with olaparib. Olaparib treatment did not affect viability, but did enhance the differentiation into mature myeloid cells. It also reduced the frequency of CD163+ (sometimes considered a marker of pro-tumor M2 macrophages) cells and induced CD80 expression. Like in the mouse model, there was a significant increase in pTBK1, PD-L1, and CSF1R levels in the macrophages, and the CSF1R+ population also expressed CD206. When olaparib treatment was started after maturation, no differences in phenotype were found in the mature myeloid cells, suggesting olaparib only affects differentiating macrophages.
Further confirmation of the data was obtained by RNAseq of the olaparib-treated macrophages. Genes associated with myeloid cell development, LPS-like inflammatory responses, and IFN signaling were significantly enriched after treatment. Proteomic analyses confirmed decreases in CD14 and CD163, as well as CD86 expression. Additionally, changes in the lipid metabolism of these cells were found, including increases in fatty acid metabolism regulation. Olaparib-treated cells were also found to have a reduction in the oxygen-consumption rate and glycolytic activity.
Macrophages that are dependent on lipid metabolism may contribute to immune suppression, and inhibition of fatty acid synthesis and SREBP1 has been shown to enhance checkpoint inhibition efficacy. Therefore, the researchers then determined whether the SREBP1 pathway regulates the olaparib-induced macrophage phenotype. By inhibiting SREBP1, the expression of PD-L1 and CSF1R induced by olaparib could be circumvented, suggesting SREBP1 was a major regulator of the olaparib-induced effects.
Since anti-CSF1R therapy has been shown to deplete tumor-promoting macrophages, the researchers hypothesized that this strategy could improve olaparib efficacy. Using the mouse model with BRCA1-deficient TNBC tumors, it was shown that olaparib inhibited tumor growth and prolonged median survival. While α-CSF1R alone had no effects, combining it with olaparib led to prolonged overall survival and complete elimination of tumors in most mice at day 35, when treatment stopped. Fewer F4/80+ and CD206+ macrophages infiltrated the tumors, and expression of CSF1R and PD-L1 was limited, changing the balance to favor antitumor macrophages over pro-tumor macrophages. Tumor cells with restored BRCA1 and two other BRCA-independent mouse tumor models did not respond to olaparib or combination treatment.
To study whether factors produced by the macrophages were able to induce tumor cell death, BRCA1-proficient TNBC cell lines were treated with conditioned media from olaparib-treated macrophages. The media induced cell death, while olaparib did not. Additionally, treatment of T cells with the media reduced viability and induced apoptosis, while olaparib enhanced T cell viability. The conditioned media also reduced glycolytic ATP and increased mitochondrial ATP in T cells.
To determine whether CD8+ T cells were required for these antitumor responses, CD8+ T cells were depleted in the mice before treatment. In this setting, the survival benefit induced by the combination treatment was significantly reduced. These data suggest that both a reduction in immunosuppressive macrophages and increases in CD8+ T cells play a role in the increased efficacy of olaparib and anti-CSF1R treatment.
Lastly, the researchers studied how the SREBP1 pathway contributes to PARPi resistance. Glucose uptake experiments in tumor-bearing animals showed that olaparib reduced the uptake of glucose by macrophages and T cells, which could be restored with treatment with α-CSF1R, supporting a metabolic switch. In addition to olaparib and α-CSF1R, when mice also received fatostatin, a fatty acid synthesis and SREBP1 inhibitor, survival improved even more. However, mice treated with olaparib and fatostatin did not display increased survival.
These final data suggest that CSF1R+ macrophages and regulation of metabolism through the SREBP1 pathway are involved in the PARPi resistance seen in a subset of patients. These results suggest that combining PARPi with anti-CSF1R or other agents targeting the inhibitory CSF1R+ macrophages is an interesting, mechanism-identified immune-targeting approach to treat patients with BRCA-mutant tumors resistant to PARPi.
Write-up by Maartje Wouters, image by Lauren Hitchings
Meet the researcher
This week, first author Anita Mehta answered our questions.

What prompted you to tackle this research question?
Triple-negative breast cancer (TNBC) is highly aggressive type of breast cancer, and PARP inhibitors (PARPi) are FDA-approved to treat TNBC patients with BRCA mutations. However, the benefits are transitory and patients on PARP inhibitor therapy eventually relapse. We have previously shown that PARP inhibitors induce an influx of white blood cells, including CD8+ T cells, into tumors, and that this influx is mediated by activation of the STING pathway in a pre-clinical model of BRCA1-deficient TNBC (Pantelidou C., Cancer Discovery, 2019). Interestingly, we observed that macrophages were actually the predominant type of infiltrating immune cells after PARPi therapy in this model. Given that tumor-associated macrophages (TAMs) generally have pro-tumor functions, we hypothesized that resistance to PARP inhibitors may be driven by suppressive macrophages.
What was the most surprising finding of this study for you?
We were most surprised by the finding that macrophages that infiltrated tumors after treatment with the PARPi olaparib displayed increased expression of antitumor markers CD80, CD86, and CD40, with concomitant increases in pro-tumor markers CD206, PD-L1, and CSF1R, indicative of anti- as well as pro-tumor tumor macrophages, which challenges the dichotomy of macrophage polarization as being either anti- or pro-tumor. To counteract the induction of immune-suppressive macrophages, we combined PARPi therapy with an antibody against the CSF1 receptor (which specifically depletes pro-tumor TAMs) in mice harboring BRCA1-deficient TNBC. This combination significantly enhanced the antitumor function of macrophages and CD8+ T cells, and extended regressions obtained with PARPi alone. The underlying mechanism of this combination still remains to be elucidated; certainly a lot of work is left to be done.
What was the coolest thing you’ve learned (about) recently outside of work?
I love cooking and always wanted to grow my own kitchen garden. During the COVID-19 pandemic, I got the opportunity to grow my own plants (tomatoes, swiss chard, kidney beans, kale, broccoli, brussel sprouts, eggplant, okra, zucchini and cilantro). It was so much fun with my 5-year-old daughter to teach her how a seed leads to a whole plant (which helped her to eat more veggies!). Now she is so curious about the basic scientific questions “How?” and “Why?” Also, it is a cool thing to know that in less than a year in the pandemic, we have learned so much about the epidemiology of this novel virus, which has led to the development of numerous vaccines in such a short time frame. This shows how spectacular and important science is. My sincere gratitude goes to all the frontline healthcare workers, scientists, and all essential workers who helped us in this difficult year.



