PD-L1 is doubling down on defense. It is well-known that cancer cells often upregulate PD-L1 as a mode of defense against the immune system, however, this new research by Gato-Cañas et al. suggests that PD-L1 may also play a direct defensive role within the tumor cell by conferring resistance to IFN-mediated cytotoxicity and cell death.
Trying to develop a melanoma cell line that secreted IFNβ while PD-L1 was silenced with a microRNA, Gato-Cañas et al. found the cells could be selected but could not survive. To explore the cause of the lethality, they surprisingly found that cells could only survive the secretion of IFNβ when PD-L1 expression was not silenced. They further noted that overexpression of PD-L1 abrogated the lethality of the IFNβ secretion/PD-L1-silencing microRNA combo. The researchers determined that cancer cells must utilize PD-L1 expression to negatively regulate IFNβ signal transduction. Intrigued by their findings, they delved deeper.
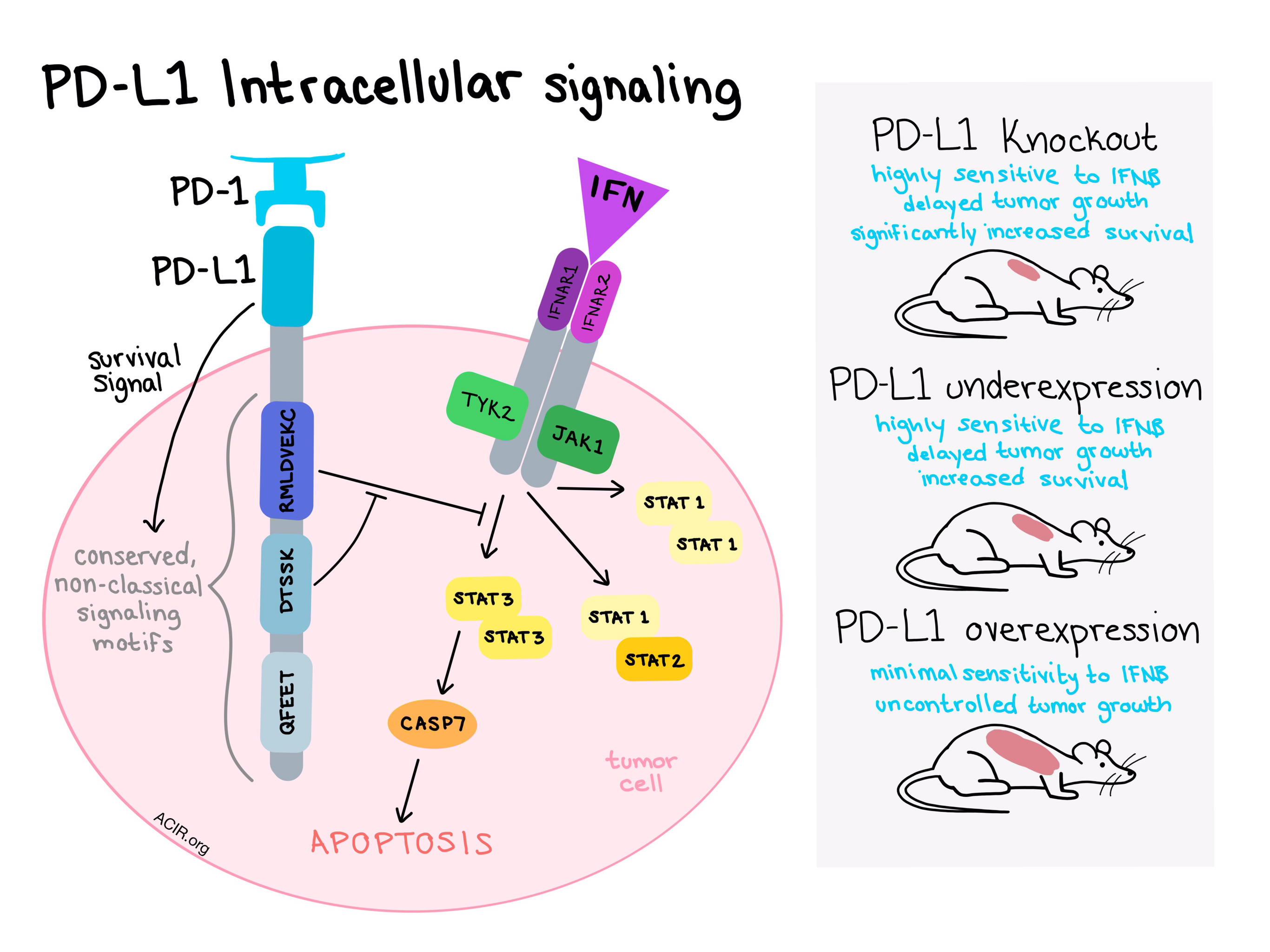
Further experiments showed that when the extracellular domain of PD-L1 engages with PD-1, it can activate an intracellular survival signal. The PD-L1 intracytoplasmic domain then transduces the signal and interferes with IFNβ cytotoxicity. When Gato-Cañas et al. analyzed the intracellular domain in search of signaling domains, however, they found none, suggesting that PD-L1 uses non-conventional signaling motifs.
In an effort to identify the mysterious signal transduction functional domains, the team compared ten mammalian PD-L1 molecules and found three phylogenetically conserved regions - “RMLDVEKC”, “DTSSK”, and “QFEET”. Experiments in which each of these motifs was deleted showed that RMLDVEKC was essential for PD-L1 to function against IFNβ cytotoxicity in the tumor cell, while DTSSK was found to be a negative regulatory motif of this activity. At least two mutations in the “DTSSK” regulatory motif were hyperactive in blocking the negative regulator, enhancing resistance to IFN-mediated cytotoxicity. Collectively, this suggested that PD-L1 intrinsic signaling within cancer cells would confer resistance to IFNs.
With the signaling domains of PD-L1 established, the next step was to determine how PD-L1 signaling crosstalks with the IFN pathway. IFNs kill cells by first engaging with their receptor, IFNAR1/IFNAR2. This recruits JAK1 and TYK2, which phosphorylate STAT1, STAT2, and STAT3, which then associate into homo- and heterodimers. IFNs also cause caspase-dependent apoptosis. By silencing various components of the known IFN pathway, Gato-Cañas et al. showed that PD-L1 interferes with the IFN pathway by inhibiting STAT3 phosphorylation and preventing STAT3 upregulation of CASP-7, stopping the signal cascade. When CASP-7 was silenced, tumor cells evaded IFNβ-mediated apoptosis. Direct blockade of PD-L1 with an anti-PD-L1 antibody sensitized murine and human cancer cells to IFNβ in vitro, indicating that cancer cells can utilize PD-1/PD-L1 engagement and subsequent signal transduction to counteract IFN toxicity.
The final step in this study was to test the IFN-resistance function of PD-L1 in vivo, independently of its role in the inhibition of T cell function. B16 tumor cells were engineered to over-or under-express PD-L1 or a PD-L1 variant lacking the cytoplasmic domain. Established tumors were then dosed directly with lentivectors expressing IFNβ. In the construct with reduced PD-L1 expression, tumor growth was delayed, and mice with the PD-L1 expression knocked out entirely showed significantly increased survival. In models that overexpressed PD-L1, tumors were protected against IFN-mediated cell death compared to controls.
Collectively, these results suggest that PD-L1 signal transduction contributes significantly to the protection of cancer cells from type I IFNs and that upregulation of PD-L1 serves as an intrinsic tumor defense mechanism against an immune attack, independent of its role in suppression of the T cell response. Blocking the IFN pathway or upregulating PD-L1 helps tumors by favoring their escape from immune attack.
by Lauren Hitchings



