A growing understanding of the gut microbiome has uncovered critical roles in regulating host immunity, metabolic health, and progression of diseases such as cancer. Recently, the composition of the microbiome has been linked to anti-PD-1 therapeutic response. Fecal microbiota transplant (FMT), in which favorable bacterial species (in this case, those conducive to anti-PD-1 responsiveness) are transferred to a host, has improved therapy outcomes in mouse models. Now, in Science, Davar and Dzustev et al. report on a first-in-human clinical trial testing the safety and efficacy of FMT with anti-PD-1 therapy in melanoma patients previously refractory to anti-PD-1 treatment.
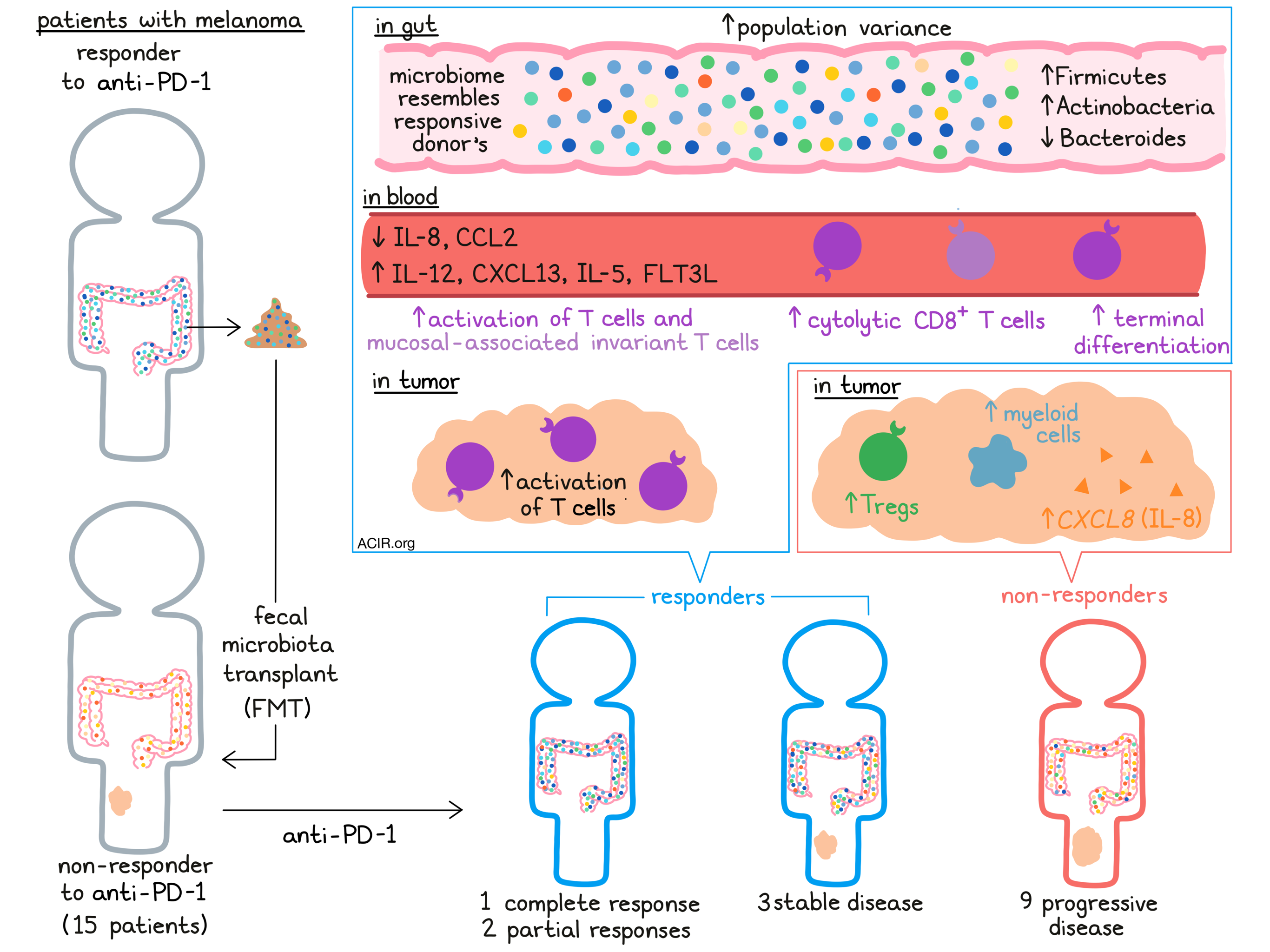
To administer FMT, Davar and Dzutsev et al. first identified potential microbiota donors. The researchers collected stool samples from metastatic melanoma patients who experienced durable remission (complete or partial responses) after previous anti-PD-1 therapy. After verifying the absence of any pathogens, donor samples were transplanted into the colon in 16 anti-PD-1-nonresponsive melanoma patients, who immediately began a new regimen of anti-PD-1 therapy (Pembrolizumab). The combination treatment (FMT + anti-PD-1) was accompanied by predominantly low-grade adverse events, induced objective responses in three patients, and supported prolonged (>12 months) stable disease in three more. Taken together, immune checkpoint blockade following FMT was feasible to deliver, safe, and effective in a subset of patients.
Assessing how effectively the transplanted bacteria incorporated into the hosts’ microbiomes, the researchers sequenced fecal samples from both recipients and donors over time. All recipients showed alterations in their gut microbiota composition, increasing population variance after FMT. Tracking composition over time, the authors observed that the recipient microbiota composition trended towards the donor microbiota profile in all anti-PD-1 responders, but only in some non-responders. In line with this result, responders had higher levels of donor bacteria-specific IgG in their circulation compared to non-responders. Looking at specific bacterial populations, the researchers found that phyla Firmicutes and Actinobacteria were enriched, while Bacteroides was reduced within responders. Interestingly, one patient who was required to take antibiotics mid-trial (due to a soft-tissue infection) showed a significant change in microbiome composition with reductions in certain beneficial species, halting antitumor benefit until a second FMT was performed. Overall, these results implied that the transplant may have been more effective in responders in whom the introduced bacteria better incorporated with pre-existing commensals and influenced host immunity.
Davar and Dzustev et al. next investigated the effects of FMT on recipient immune responses more comprehensively, collecting PBMCs for flow cytometric analysis. They found that responders had higher proportions of circulating CD56+CD8+ T cells, noted for their cytolytic capacity, compared to non-responders. Furthermore, responding patients’ CD8+ T cells upregulated TIGIT, Tbet, and LAG3 after FMT, had a reduced proportion of naive T cells, and were enriched in a terminally differentiated effector-memory phenotype. Broadly, FMT appeared to induce an activated T cell phenotype in the blood of patients who went on to respond to anti-PD-1 therapy. In agreement with these results, mucosal-associated invariant T cells also expressed higher activation markers (e.g., granzyme B) in responders versus non-responders.
FMT also displayed striking effects within tumors, from which the researchers isolated CD45+ immune cells for analysis by scRNAseq. Tumors from non-responding patients contained a greater proportion of myeloid cells and Tregs compared to responders, indicative of immune suppression. Particularly, myeloid cells in the non-responding patients expressed high levels of CXCL8, which encodes the immunosuppressive cytokine IL-8. The intratumoral T cell compartment matched observations made in the blood; tumor-infiltrating T cells in responders similarly displayed an activated profile with upregulation of markers including GZMK.
To further examine the systemic effects of FMT, the authors next performed multi-omic analyses on patient serum samples. Compared to non-responders, responders demonstrated considerable changes in serum cytokines and chemokines, lipidomic profiles, and circulating metabolites after treatment. Notably, responders after FMT had reduced IL-8 and CCL2, and increased IL-12, CXCL13, IL-5, and FLT3L – altogether a favorable cytokine profile for antitumor immunity. The observation of decreased IL-8 in circulation complemented the intratumoral myeloid analysis in these patients. Responding patients also increased serum triacylglycerols (with decreases in mono- and diacylglycerols), as well as bile acids and products of bacterial catabolism compared non-responding patients. Taken together, these data indicate that FMT induced comprehensive, systemic effects in responding patients.
Finally, to discern relationships among the extensive collected data, the authors constructed a computational model to link bacterial populations, cytokine/chemokine profiles, immune cells, metabolites and lipids. They observed that nodes representing microbiota were highly interconnected with the others, suggesting a central role of the microbiome in regulating these systemic effects. Additionally, the researchers derived that the bacterial species found to increase and decrease in responding patients correlated negatively and positively with IL-8, respectively, further highlighting IL-8 as an important regulator of antitumor responses.
Overall, Davar and Dzustev et al. demonstrated the direct clinical relevance of the microbiome to therapeutic outcomes in cancer. A single fecal transplant altered the microbiome of recipients, in turn modulating both systemic and intratumoral immune responses and sensitizing previously refractory patients to anti-PD-1 therapy. Further understanding of the mechanisms underlying these relationships and the precise identification of favorable microbial species or combinations of species, as well as additional data from clinical trials, will continue to advance these therapies – and likely incorporate microbiome profiling in next-generation precision cancer medicine.
Write-up by Alex Najibi, image by Lauren Hitchings



