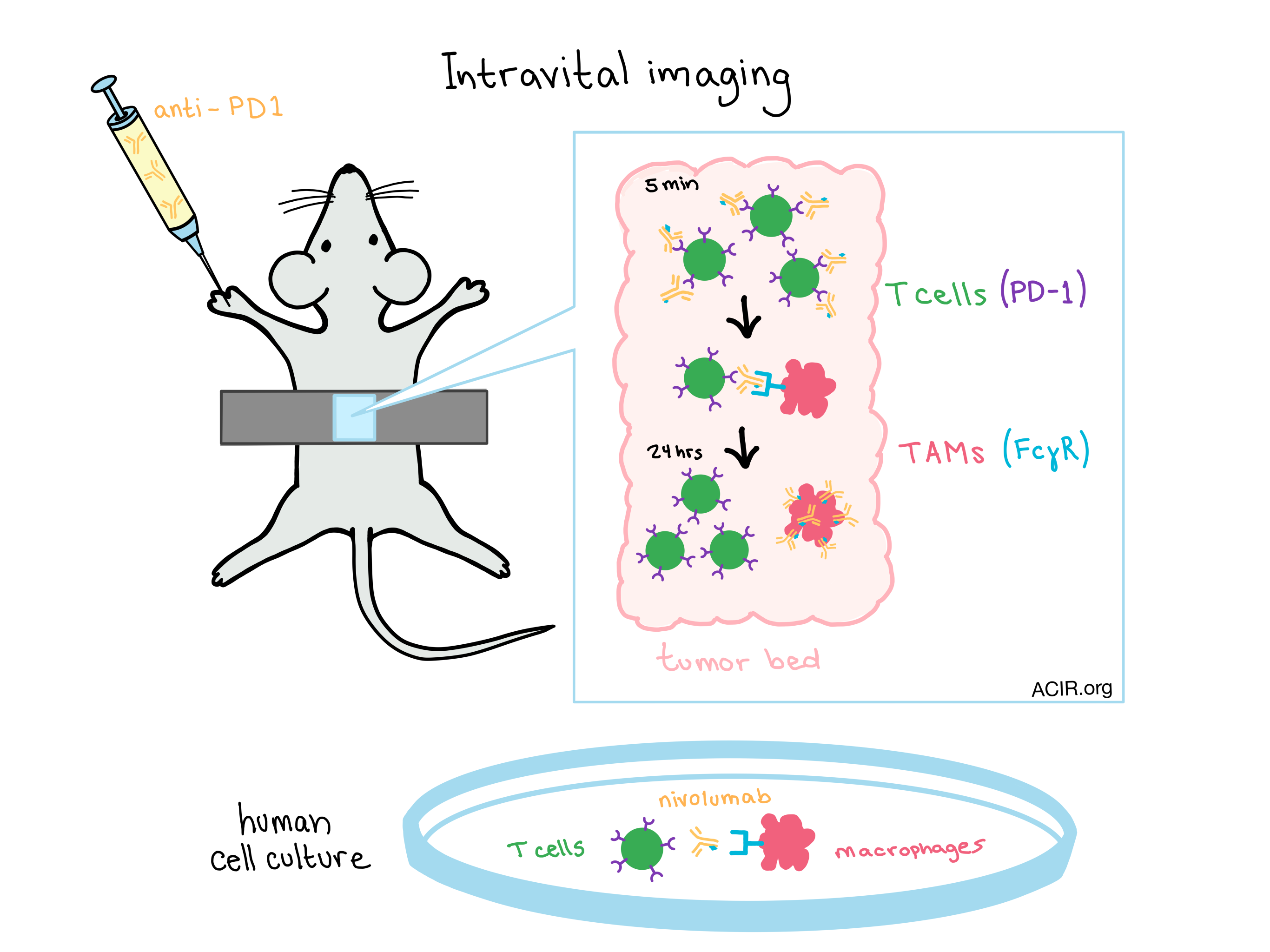
In order to fully understand the mechanism of PD-1 checkpoint blockade therapy and potentially reduce the number of nonresponders, it is important to understand exactly what is happening in vivo. For that reason, Arlauckas et al. used intravital imaging to follow a fluorescently-labeled anti-PD-1 monoclonal antibody (mAb) in real time at a subcellular resolution and track its interactions with CD8+ T cells, tumor cells, and tumor-associated macrophages (TAMs), each labeled with a unique fluorescent marker.
Intravital imaging revealed the following:
The anti-PD-1 signal began to accumulate in the tumor within minutes of injection. Maximum accumulation occurred after 24 hours.
The antibody rapidly perfused tumor blood vessels and gradually migrated into the tumor bed.
Initially, the anti-PD-1 drug bound to the surface of CD8+ T cells in the tumor microenvironment.
Later time points showed that TAMs, which are stationary, had collected most of the anti-PD-1 through physical interactions with neighboring motile T cells. At this later point, anti-PD-1 was no longer associated with T cells.
The pattern of anti-PD-1 theft by TAMs occurs even when the frequency of TAMs is quite low.
Further data suggest that macrophages eventually degrade the antibody.
Mechanism of anti-PD-1 transfer: In an effort to further explore the mechanism by which anti-PD-1 is transferred from PD-1+ T cells to PD-1- macrophages, the researchers designed an in vitro coculture system of the two cell types. The researchers again saw that macrophages did not capture free anti-PD-1 in substantial quantities early after drug administration, but then withdrew drug bound to T cell surfaces. Additional testing revealed that transfer could not be attributed solely to phagocytosis of cell debris or trogocytosis, and that loss of anti-PD-1 from T cells was not due to receptor internalization.
Suspecting that inhibitory Fc receptors played a role in how TAMs pilfer anti-PD-1 from nearby T cells, the researchers added a Fcɣ receptor (FcɣR) blocking antibody to the coculture. Confirming their hypothesis, the addition successfully diminished the transfer of the anti-PD-1 from T cells to macrophages. As FcɣR binding is known to depend on the presence of glycan structures on the antibody, the researchers used an enzyme to strip the glycan away. Flow cytometry revealed that the deglycosylated anti-PD1 still efficiently bound to PD-1+ T cells, but live cell imaging showed diminished transfer of the drug to macrophages.
Therapeutic relevance: When mice were treated with anti-PD-1 in combination with FcɣR blocking antibodies, intravital imaging showed that anti-PD-1 mAbs remained bound to CD8+ T cells in the tumor bed for a substantially longer period of time. Furthermore, the blockade of FcɣR interactions completely eliminated the fraction of anti-PD-1 nonresponders usually seen in MC38-bearing mice, and all mice that received the combination treatment showed complete tumor rejection.
Similar glycosylation patterns between anti-PD-1 in mice, and the anti-PD-1 drug nivolumab for humans predict that FcɣR-mediated antibody transfer will be relevant in humans. A co-culture of nivolumab-labeled human CD8+ T cells with human macrophages showed the same pattern of drug transfer as the mouse model, and the use of FcɣR blocking antibodies inhibited the transfer. The success of the FcɣR blockade in mice suggests that human immunotherapies utilizing anti-PD-1 blockade could be improved by blocking FcɣR interactions, either through the use of an Fc block, deglycosylation of the therapeutic antibodies, or direct targeting of TAMs.
by Lauren Hitchings



