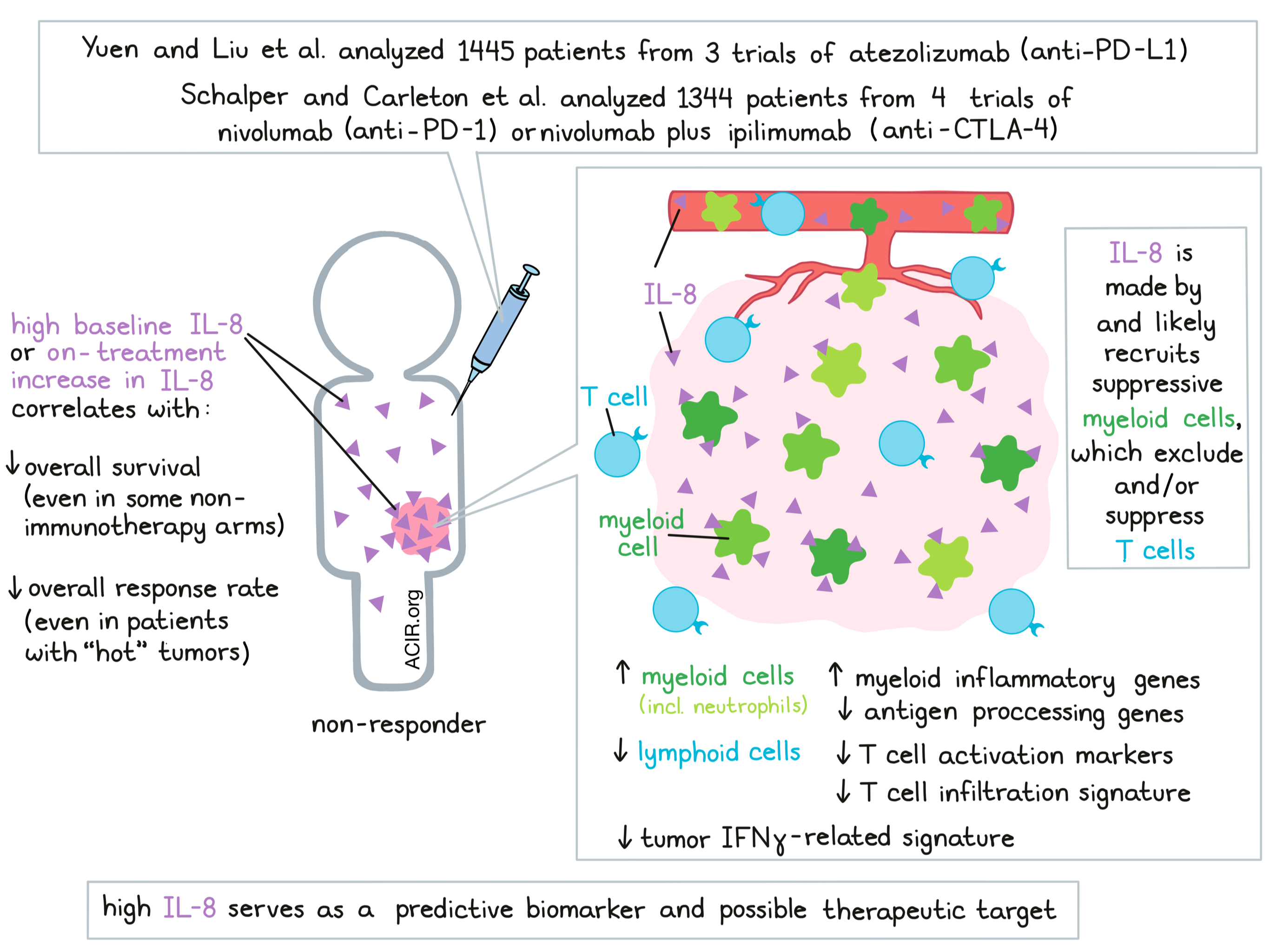
When it comes to improving cancer immunotherapy, finding strong biomarkers and finding novel targets are both key goals. In two papers recently published back-to-back in Nature Medicine, Yuen and Liu et al. and Schalper and Carleton et al. identify IL-8 (also known as CXCL8) both as a predictive biomarker of reduced clinical benefit from PD-1/PD-L1-targeted checkpoint blockade, and as a potential therapeutic target to improve the efficacy of checkpoint blockade.
IL-8 is a proinflammatory chemokine expressed in many human tumors that attracts myeloid cells and neutrophils, and may support tumor growth through a variety of mechanisms. Following up on prior evidence from smaller studies that showed IL-8 to be associated with reduced response to PD-1/PD-L1 checkpoint blockade, Yuen and Liu et al. analyzed data on plasma (p)IL-8 and IL8 gene expression in peripheral blood mononuclear cells (PBMCs) and tumors from 1445 patients across three clinical trials of atezolizumab (anti-PD-L1): IMvigor210 for metastatic urothelial cancer (mUC), IMvigor211 for mUC, and IMmotion150 for metastatic renal cell carcinoma (mRCC). Similarly, Schalper and Carleton et al. evaluated baseline serum IL-8 levels in samples from 1344 patients treated with nivolumab (anti-PD-1) monotherapy or nivolumab plus ipilimumab (anti-CTLA-4) in four clinical trials: CheckMate 067 for melanoma, CheckMate 017 for squamous NSCLC, CheckMate 057 for nonsquamous NSCLC, and CheckMate 025 for RCC.
Beginning with data from IMvigor210, Yuen and Liu et al. found that baseline pIL-8 was not correlated with typical markers of a “hot” tumor microenvironment, like an effector T signature, TMB, tumor PD-1 and PD-L1 expression, a TGFβ response signature, or a TIDE signature. In certain patient groups, however, high baseline pIL-8 did correlate with worse overall survival (OS) and lower overall response rate (ORR). This observation held true even in patients whose tumors showed high baselines effector T cell signatures that represent a “hot” TME. Extending their analysis, Yuen and Liu et al. found that in the IMvigor211 trial, patients with high pIL-8 also showed worse survival in both the atezolizumab and control chemotherapy arms, suggesting that high pIL-8 was broadly prognostic in mUC. In the IMmotion150 trial, high pIL-8 was associated with worse OS in atezolizumab arm, but was not statistically significant in the atezolizumab plus bevacizumab arm nor the sunitinib arm.
Performing a similar retrospective analysis, Schalper and Carleton et al. stratified patients based on high or low baseline serum IL-8 and found that high serum IL-8 was associated with shorter OS across treatments and tumor types. The strongest detrimental effect of high IL-8 was observed in patients with melanoma treated with nivolumab plus ipilimumab. It was also associated with lower ORR and poorer survival across most individual treatment arms investigated, including non-immunotherapy arms.
To evaluate whether on-treatment changes in IL-8 affected outcomes, Yuen and Liu et al. evaluated pIL-8 levels 6 weeks after treatment. In IMvigor210, an on-treatment increase in IL-8 was associated with worse OS and ORR in both atezolizumab- and chemotherapy-treated patients. In the IMvigor211 trials, the same effect was observed only in the atezolizumab treatment arm.
Next, Yuen and Liu et al. used single-cell RNAseq to evaluate IL8 expression in PBMCs from 5 responders and 5 nonresponders from the IMvigor210 trial and found that nonresponders had a larger portion of IL-8-producing cells and higher overall expression of IL8. IL8 expression was higher in myeloid clusters than lymphoid clusters, and cells expressing high levels of IL8 also showed high expression of myeloid inflammatory genes and lower expression of genes related to antigen-presenting machinery. Like serum pIL-8, high IL8 gene expression in PBMCs was significantly associated with worse OS in both arms of the IMvigor210 trial, and in the atezolimumab arms of the IMvigor211 and IMmotion150 trials.
To further investigate the role of IL-8 in the tumor microenvironment, Yuen and Liu et al. compared fresh tumors and peripheral blood samples from four patients with RCC. Using scRNAseq and UMAP visualization, the researchers identified 11 phenotypic clusters of immune cells. Compared to blood, tumors were enriched for myeloid populations, which had higher IL8 expression than lymphoid populations. Within myeloid populations, IL8 expression was most pronounced in tumor-infiltrating macrophages (particularly M1 macrophages), and less pronounced in myeloid cells from peripheral blood. Within tumor-infiltrating myeloid cells, cells with high IL8 expression showed increased expression of proinflammatory genes, reduced expression of genes involved in antigen processing and presentation, and reduced expression of IFN-inducible genes.
Yuen and Liu et al. found that IL8 expression correlated with higher neutrophils in tumors in both the IMvigor210 and IMmotion150 trials. In patients with low pIL-8, CD8+ T cells had high expression of T cell activation markers and other T cell-associated genes. Similarly, Schalper and Carleton et al. found that circulating IL-8 correlated with blood neutrophil and monocyte counts. Local expression of IL-8 in tumors positively correlated with infiltration of neutrophils and tolerogenic myeloid cells, and negatively correlated with transcripts related to T cell infiltration and IFNγ.
Based on their results, both Yuen and Liu et al. and Schalper and Carleton et al. suggest that IL-8 likely recruits immunosuppressive myeloid cells like neutrophils and macrophages to patients’ tumors, resulting in tumor environments that exclude T cells and/or suppress T cell activation. Therefore, IL-8 is likely representative of a suppressive TME, which would explain why high baseline IL-8 serves as an independent biomarker that predicts shorter survival and reduced clinical benefit from PD-1/PD-L1 checkpoint blockade.
In the clinic, IL-8 has the potential to be particularly useful as a biomarker to identify patients who are unlikely to respond to PD-1/PD-L1 checkpoint blockade, as it can easily be measured from blood samples; Schalper and Carleton et al. even proposed a clinically meaningful cutoff level for baseline serum IL-8 of 24 pg/ml. IL-8, or its receptors, CXCR1 and CXCR2, could also be considered as potential therapeutic targets to enhance responses to checkpoint blockade.
by Lauren Hitchings



