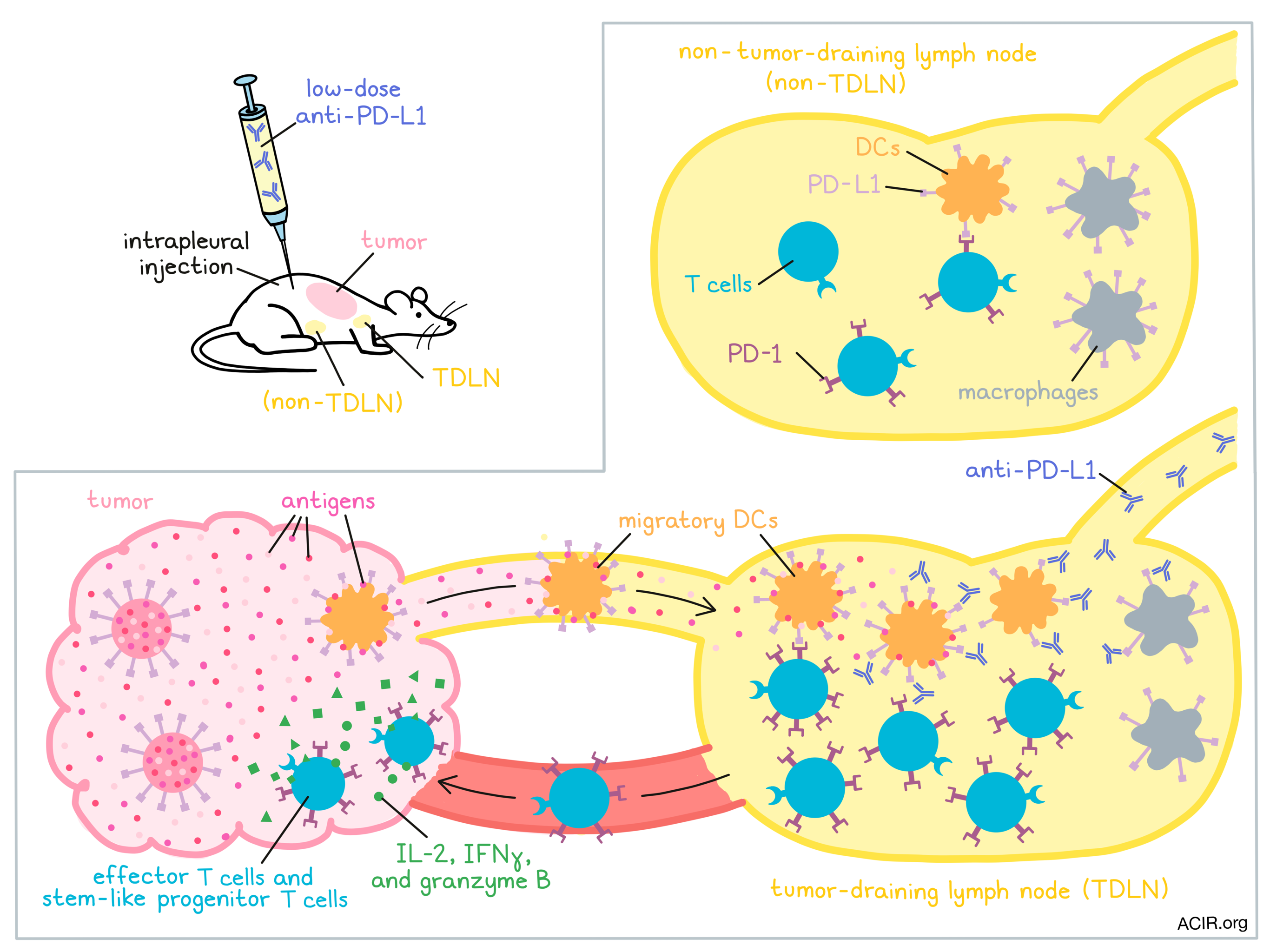
Targeting the PD-1/PD-L1 axis is a common immunotherapeutic strategy that works well in some, but not all patients with cancer. Typically, researchers have looked at interactions between PD-1 and PD-L1 within tumors to explain the mechanism behind this treatment; however, this has never fully explained the efficacy, or lack thereof, of PD-1/PD-L1 blockade. Recently, Dammeijer and van Gulijk et al. investigated whether interactions within tumor-draining lymph nodes (TDLNs) might play a role, and their results were published in Cancer Cell.
To begin, Dammeijer and van Gulijk et al. analyzed antigen-specific T cells in the lymph nodes (LNs) of mice bearing OVA+ AE17 mesothelioma tumors. Compared to in non-TDLNs, OVA-targeted T cells in TDLNs were more abundant and more proliferative, and expressed PD-1 more often and at higher levels. Similar results were found in multiple other tumor types. However, in the poorly immunogenic KPC3 pancreatic tumor model, fewer PD-1+ T cells were found in TDLNs. The addition of OVA expression to KPC3 cells, however, induced PD-1+ T cells in the TDLN, suggesting that PD-1 expression on T cells in the TDLN is related to tumor immunogenicity.
To better track the T cell response to tumor antigens, the researchers infused naive OT-I and OT-II cells into mice with OVA-expressing tumors. The tumor-specific T cells initially migrated to secondary lymphoid organs. Upon homing to the TDLN, they upregulated PD-1 and proliferated, suggesting early activation. Eventually, these cells moved into the blood, and ultimately accumulated in tumors, where they expressed IL-2, IFNγ, and granzyme B, suggesting an effector T cell phenotype.
Having established the patterns of PD-1 expression on T cells, the researchers next sought to characterize the expression of PD-L1. Investigating PD-L1 on myeloid cells, the researchers found that in the TDLN (versus in non-TDLNs) PD-L1 expression was upregulated on macrophages (subscapular sinus macrophages (SSMs) and macrophages lining the medullary sinuses, (MSMs)) and dendritic cells (CD11b+ DCs, cDC1s, and cDC2s), but not granulocytes. PD-L1 expression was particularly high on migratory cDC2s in the TDLN.
In order to study the PD-1/PD-L1 axis specifically in TDLNs, the researchers developed a strategy in which they injected a low dose of anti-PD-L1 antibodies intrapleurally. The antibodies then drained directly to the mediastinal LNs, which are the TDLNs for intraperitoneal tumors. Because of the low dose though, the antibodies did not drain into circulation or into the tumors. Also because of the low dose, not all of the PD-L1 molecules in the TDLN were blocked. Surprisingly, this treatment still mediated control of AC29 mesothelioma and MC38 colon carcinoma tumors and provided a survival benefit in mice. While it did not induce the same level of proliferation or T cell infiltration into tumors as high-dose systemic anti-PD-L1 therapy, it did induce infiltration of KLRG-1+ effector T cells and of CD4+ and CD8+ T cells with a stem-like progenitor phenotype.
When the same experiment was repeated in the aggressive OVA+ AE17 mesothelioma model, the researchers observed that anti-PD-L1 induced proliferation and accumulation of antigen-specific T cells in TDLNs, but not non-TDLNs. Progenitor T cells were found in TDLN within 24 hours of anti-PD-L1 treatment, in blood at 72 hours, and in the tumor at 144 hours. In an ex vivo stimulation assay of T cells in their natural TME, stimulation of T cells with OVA peptide increased CD8+ T cell activation, while stimulation with anti-PD-L1 led to upregulation of PD-1. In samples taken from mice treated with TDLN-targeted anti-PD-L1, TIL activity was enhanced and PD-1 levels were reduced. Together these results suggested that PD-L1 blockade in the TDLN reduced T cell suppression.
To prove that T cells from the TDLN contributed to antitumor immunity, Dammeijer and van Gulijk et al. used the S1P receptor agonist FTY720 to abrogate T cell egress from lymphoid organs. This prevented the antitumor efficacy of anti-PD-L1 and tumor infiltration by CD4+, CD8+, and stem-like progenitor T cells.
To evaluate how different cell types contribute to PD-L1-mediated immunosuppression, the researchers depleted TDLN macrophages. This did little to affect the antitumor efficacy of anti-PD-L1 therapy, suggesting that PD-L1 on cDCs was more influential. Supporting this, the researchers observed colocalization between CD8+ T cells and PD-L1+ DCs, but not between CD8+ T cells and macrophages in tumors.
Having shown that PD-L1 on cDC2s mediated suppression of T cells in the TDLN, and that these interactions contributed to the efficacy of anti-PD-L1 immunotherapy, Dammeijer and van Gulijk et al. investigated whether the same observations would hold true in patients. To evaluate the contribution of the TDLN, the researchers looked at patients with stage II melanoma who had received no systemic treatments. Following surgical resection of tumors, some of these patients had early disease recurrence, while others remained disease-free long term. While patients with early recurrence also had other primary tumor characteristics that served as prognostic indicators for recurrence (tumor depth, ulceration, etc.) they also had higher PD-1/PD-L1 interaction densities within TDLNs compared to patients who remained disease-free long-term, and this increased PD-1/PD-L1 interaction density served as an independent prognostic factor. In tumor samples, the researchers observed contact between CD8+ T cells and cD11c+ DCs, but rarely between T cells and macrophages. Interestingly, while PD-L1 levels were higher in tumors than in the TDLNs, PD-1/PD-L1 interactions were scarce within tumor tissue.
Overall, Dammeijer and van Gulijk et al. present a model in which tumor antigens and migratory DCs make their way to TDLNs to activate T cells, and that interactions between PD-L1 on DCs and PD-1 on T cells in the TDLN critically influence the antitumor efficacy of the T cell response. In patients, the density of interactions between PD-1 and PD-L1 in the TDLN could help to identify patient groups with a high risk of disease recurrence. This research also highlights TDLNs as hubs of antitumor immunity, and challenges the notion that PD-1/PD-L1 blockade acts mainly at the tumor site. This new understanding may also support the use of neoadjuvant PD-1/PD-L1 checkpoint blockade, as it may be more effective to begin treatment before removal of the tumor (as the antigen source) and the TDLN.
Lauren Hitchings
Meet the researcher
This week, first co-author Floris Dammeijer answered our questions.
What prompted you to tackle this research question?
PD-1/PD-L1 checkpoint blockade immunotherapy has revolutionized the treatment of cancer, however, still a majority of patients and cancer types does not respond durably. PD-L1 expression in the tumor has been investigated extensively as a biomarker, but has been found to suboptimally stratify patients; there are PD-L1 non-expressing patients demonstrating response to immunotherapy and vice versa. Therefore, we turned to non-tumor sites, including tumor-draining lymph nodes (TDLNs), as these are constantly exposed to tumor-derived molecules and involved in immune response generation. We hypothesized that PD-1/PD-L1 interactions in TDLNs suppress the induction and propagation of effective antitumor immunity, and that antibodies blocking these checkpoints work in part through TDLNs.
What was the most surprising finding of this study for you?
When we targeted TDLNs specifically, but suboptimally, with a 100-fold lower dose of PD-L1-blocking antibody, we were astonished that we could generate effective immune responses in the tumor. These findings in several mouse models indicated for the first time that PD-L1 blockade in TDLNs alone is sufficient to generate systemic immunity. Furthermore, our findings in stage II (non-metastatic) melanoma patients sparked our interest, as we could identify high levels of PD-1/PD-L1 interactions in TDLNs to be prognostic of early, distant disease recurrence. Besides being in line with our murine data, this assay potentially identifies a novel high-risk patient subgroup that could benefit from adjuvant PD-1/PD-L1 blockade immunotherapy.
What was the coolest thing you’ve learned (about) recently outside of work?
Recently, I visited a coastal part of the Netherlands with one of my co-authors in search of wild oysters and learned a lot about these amazing, yet tasty creatures. Did you know they can filter up to a bathtub of water every day and can age up to 30 years? When eating a wild oyster, check if it is firmly closed, does not contain any sand or crabs, and sheds its own water; now they are safe to shuck and eat. If you’re unsure how to shuck an oyster and want to ever wield a pipet again, wear special gloves to protect your hands. Add vinaigrette (equal parts white- and red-wine vinegar, diced shallot, salt, pepper, sugar) or incorporate into a seafood pasta!




