The gut microbiome has recently emerged as a critical regulator of antitumor immunity. Nearby, the liver is supplied by blood largely from the intestines, meaning that it is directly exposed to the runoff of metabolites and other products of the microbiome. Noting this direct physical relationship, Ma et al. set out to determine whether changes in the microbiome might affect hepatic immune cells and antitumor immunity in the liver.
To begin, the researchers induced spontaneous hepatocellular carcinoma (HCC) in mice and treated them with an antibiotic cocktail (ABX) to deplete commensal gut bacteria. Surprisingly, they found fewer and smaller HCC tumors in mice treated with the antibiotics. Similarly, in liver metastasis models (subcutaneous EL4 thymoma and intrasplenic tumor injection of B16 melanoma), fewer spontaneous metastases formed in the liver when mice were treated with antibiotics, while no effects were observed on primary tumors located outside of the liver, nor on the formation of metastases in other locations, indicating a liver-specific antitumor effect.
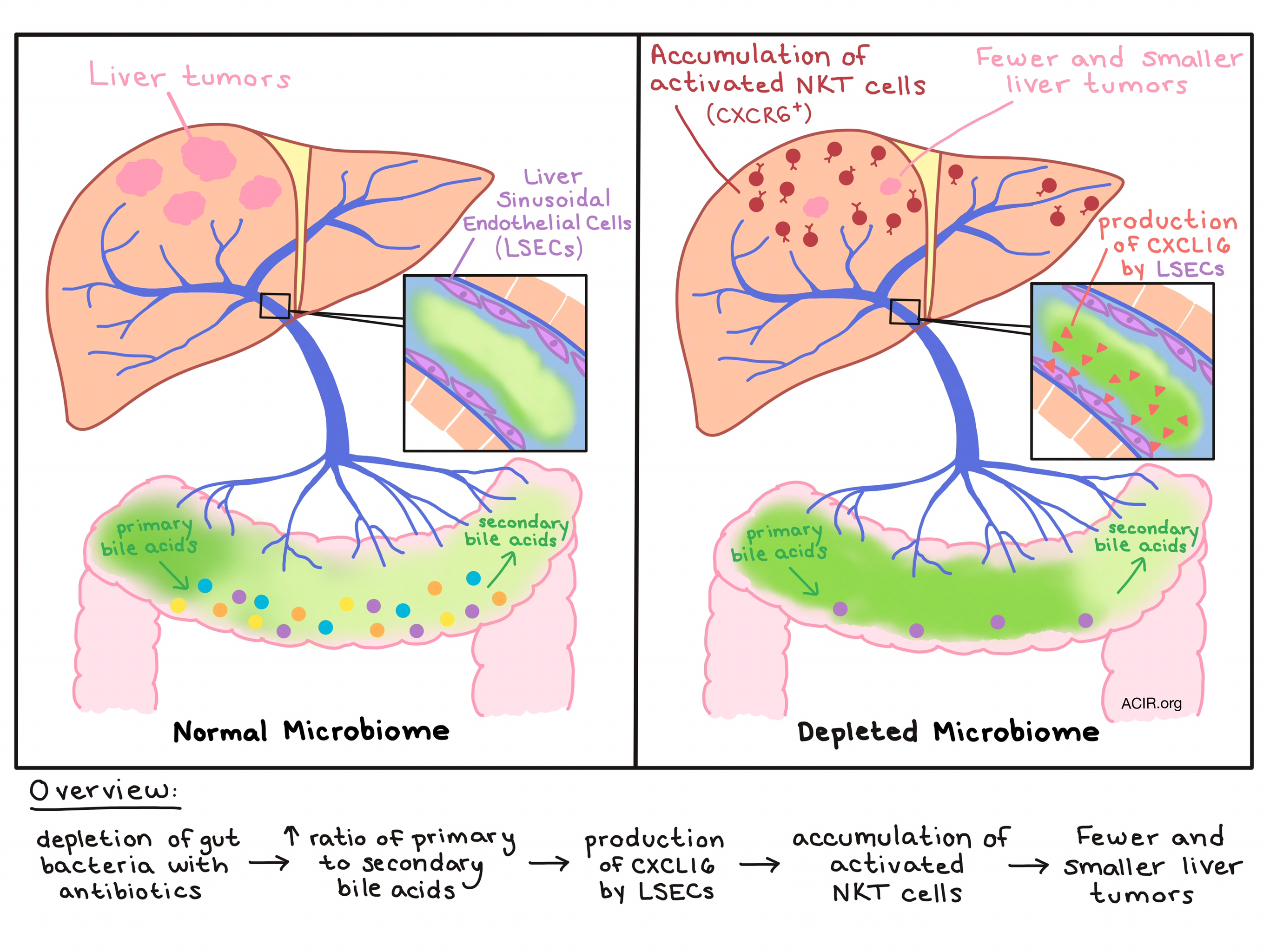
Exploring which immune cell subsets might be behind the observed antitumor effect, Ma et al. noted prominent expansion and accumulation of hepatic natural killer T (NKT) cells in the livers of ABX-treated mice, and that those NKT cells were highly activated and produced higher levels of IFNγ, both in tumor-bearing and tumor-free mice. Antibody depletion and gene knockout experiments in tumor-bearing mice showed that NKT cells were the primary mediators of tumor inhibition associated with ABX treatment.
Nearly all hepatic NKT cells express the receptor CXCR6, which interacts only with the ligand CXCL16, so unsurprisingly, Ma et al. found that ABX treatment increased mRNA levels of Cxcl16. Previously reported to be a major source of CXCL16, liver sinusoidal endothelial cells (LSECs) were found to upregulate CXCL16 production in ABX-treated mice, thus recruiting NKT cells to the liver.
Based on a known role for the cell surface-bound form of CXCL16 in lipid metabolism, Ma et al. suspected a link between CXCL16 upregulation by LSECs and bile acid metabolism mediated by commensal gut bacteria. To prove this connection, the team used a bile acid sequestrant to reduce bile acid levels in the liver and saw increased NKT cell accumulation. Digging deeper into which bile acids were at play, the researchers found that ABX treatment increased primary acids, while reducing secondary bile acids, consistent with the understanding that commensal gut bacteria convert primary bile acids into secondary bile acids. In vitro, secondary bile acids were found to suppress expression of Cxcl16 mRNA by LSECs, while primary bile acid induced expression of Cxcl16. In vivo, oral gavage of secondary bile acids reversed the ABX-mediated accumulation of NKT cells and antitumor effect in the liver, while primary bile acids had the opposite effect - further enhancing NKT cell accumulation and tumor inhibition.
By treating mice with individual antibiotics rather than a cocktail, the research team was able to identify gram positive bacteria of the Clostridium genus as having a negative effect on hepatic NKT cell accumulation and liver tumor inhibition, and reconstitution with C. scindens rapidly reduced the effects of ABX treatment. Depleting these bile-acid converting commensal gut bacteria raises the ratio of primary to secondary bile acids, inducing production of CXCL16 by LSECs in the liver. Increased CXCL16 production leads to the accumulation of activated CXCR6+ NKT cells in the liver, which can exert an antitumor effect on primary liver tumors and liver metastases in mice.
Ma et al. extended their outlook to human samples to determine whether their findings in mice might translate. Similar to what they saw in mice, primary bile acids induced CXCL16 mRNA expression in SK-HEP1 cells, representative of LSECs. Analysis of primary/secondary bile acid ratio and CXCL16 mRNA levels in data from a prior cohort of patients suggested that the opposing effects of primary and secondary bile acids on CXCL16 expression hold true in humans. In addition to providing insight into the connection between the microbiome, the immune system, and liver cancer, the results of this study may influence the design of future immunotherapies related to the treatment of liver cancer or liver metastases.
by Lauren Hitchings



