The PD-1 axis plays an important role in regulating the activation of T cells, and blockade of the PD-1 axis has had profound clinical significance over the past decade. Although multiple studies have investigated the impact of PD-1 signaling during the early stages of an immune response and on the rescue of “exhausted” T cells, its exact role in affecting differentiation towards long-term memory phenotypes is not clear. To this end, Pauken, Godec, and Odorizzi et al. set out to uncover the specific role of the PD-1 axis in the formation of memory T cell responses; the results of their investigation were recently published in Cell Reports.
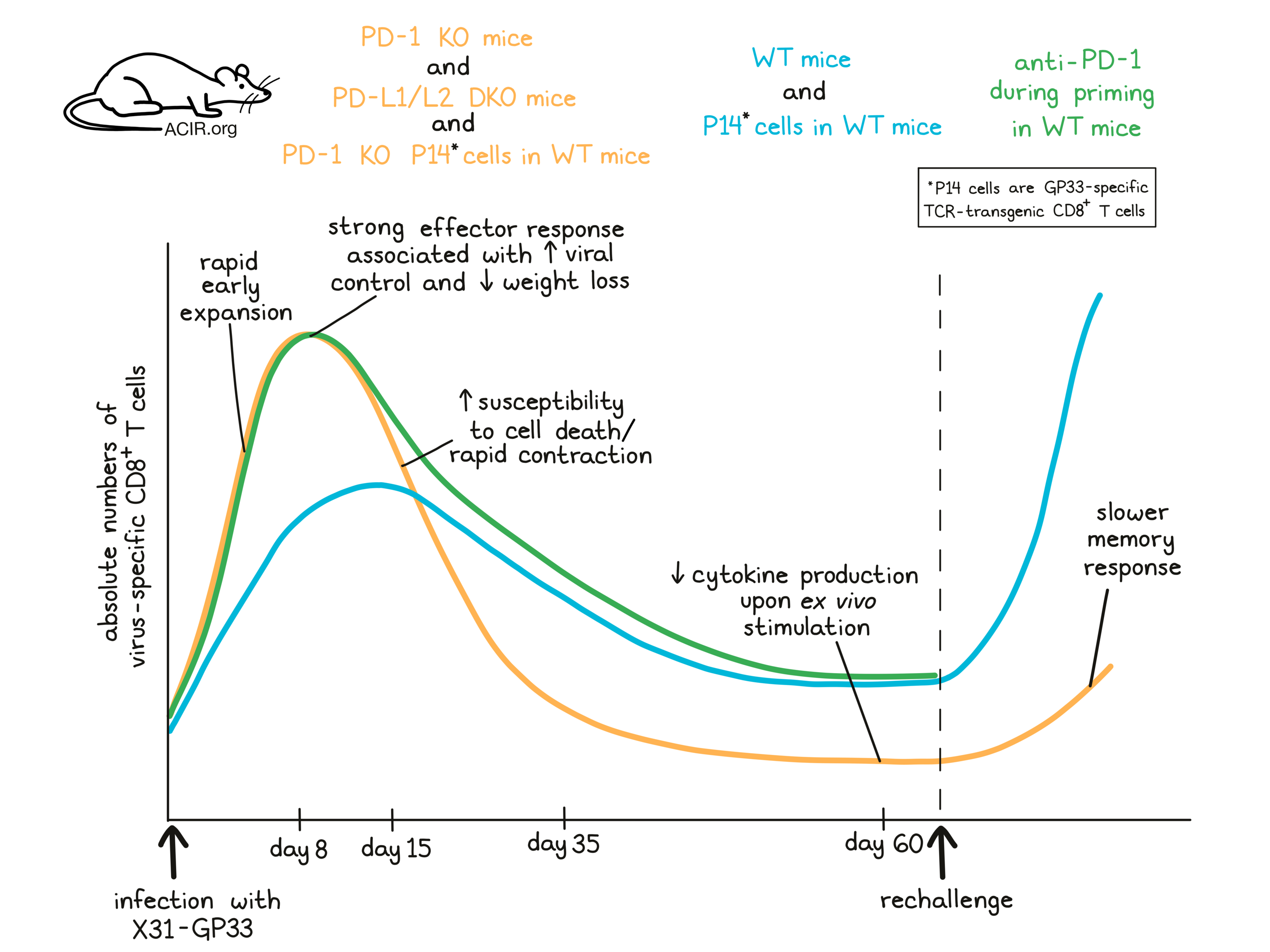
To begin, Pauken, Godec, and Odorizzi et al. showed that in wild-type (WT) mice infected with the acute influenza strain X31 carrying the GP3333-41 epitope from LCMV (X31-GP33), PD-L1 and PD-L2 became upregulated on antigen-presenting cells, while PD-1 was rapidly upregulated on CD4+ and CD8+ T cells in the lung, with high levels maintained through day 12. Next, the researchers infected mice lacking either PD-1 (PD-1 knockout [KO]) or both PD-L1 and PD-L2 (PD-L1/L2 double knockout [DKO]) with X31-GP33 and followed virus-specific CD8+ T cells from days 8 through 10 post infection (p.i.). While virus-specific CD8+ T cells in the lungs were observed at similar frequencies in WT and PD-L1/L2 DKO mice, absolute numbers were higher in PD-L1/L2 DKO mice, suggesting that the lack of PD-1 axis signaling increased the magnitude of CD8+ T cell responses in the lung. Further, PD-1 KO and PD-L1/L2 DKO mice lost less weight than WT mice, and the increased number of virus-specific CD8+ T cells correlated with reduced viral loads, suggesting enhanced immune control.
To better understand the mechanism behind the increased number of virus-specific CD8+ T cells, Pauken, Godec, and Odorizzi et al. performed gene set enrichment analysis at day 8 p.i. and found that while pathways related to cell division were enriched in virus-specific CD8+ T cells from WT mice, cells from PD-L1/L2 DKO mice appeared to be exiting the cell cycle. Further, a lower frequency of CD8+ T cells appeared to be proliferating (KI-67+) in the lungs of PD-L1/L2 DKO mice, while a higher frequency appeared to be proliferating in the spleen. A higher frequency of virus-specific CD8+ T in the lungs of DKO mice also showed active caspase staining, suggesting that these cells were more susceptible to death. Together, this suggested that virus-specific CD8+ T cells in mice lacking a PD-1 pathway underwent early and excessive proliferation, and that those in the lung had already entered the contraction phase of the response at day 8 p.i.
Having observed that effector T cell responses were altered in mice lacking a PD-1 pathway, Pauken, Godec, and Odorizzi et al. hypothesized that these early changes might affect the formation of CD8+ memory T cells, and extended their analysis to longer timepoints. The researchers analyzed CD8+ T cells specific for GP33 and two natural flu epitopes and found that from day 8 to 10 p.i., CD8+ T cells specific for all three epitopes were present at higher numbers in PD-L1/L2 DKO mice compared to WT. However, at day 15, responses to GP33 and one of the flu epitopes showed greater contraction in PD-L1/L2 DKO mice. By day 35, CD8+ T cells specific for all 3 epitopes were lower in PD-L1/L2 DKO mice compared to WT, and by day 60, both the frequency and absolute number of virus-specific CD8+ T cells were lower in PD-L1/L2 DKO mice, suggesting that without PD-1 signaling, initial CD8+ T cell responses to acute viral infection were greater, but fizzled out more rapidly, while in WT mice, the response was less intense, but more sustained. Virus-specific CD8+ T cells taken from PD-L1/L2 DKO mice at day 60 showed lower cytokine production upon ex vivo stimulation compared to WT, further suggesting a qualitative defect in the memory response as well.
To test whether the differences in virus-specific CD8+ T cells would affect secondary responses, mice were rechallenged with another influenza strain also expressing GP33, which would require a T cell response in order to be eliminated. Compared to WT mice, PD-L1/PD-L2 DKO mice had higher amounts of virus in their lungs and lost more weight during secondary infection. They also had lower frequencies and lower total numbers of influenza-specific and cytokine-producing CD8+ T cells in the lungs 3.5 days post rechallenge. These results suggested that a lack of PD-1 pathway signaling impaired and delayed protective memory responses, leading to more severe secondary infections.
To determine whether the impaired CD8+ memory T cell responses in PD-1 pathway-deficient mice were caused by cell-intrinsic or cell-extrinsic factors, the researchers adoptively transferred either WT or PD-1 KO GP33-specific TCR-transgenic CD8+ T cells (P14) into WT mice, and then infected the mice with X31-GP33. Like in the complete knockout models, PD-1 KO P14 cells expanded early, exhibited enhanced effector responses, were more prone to cell death in the lungs, underwent more rapid contraction, were less abundant in the lung and spleen after 3 months, and responded less potently to ex vivo stimulation at 3 months compared to WT P14 cells. Upon rechallenge, PD-1 KO P14 memory cells were outperformed by WT P14 memory cells, which protected mice more quickly and more thoroughly. These results suggested that the role of PD-1 in affecting memory functions was intrinsic to the cell. Similar results were observed when mice were infected with two other model respiratory viruses.
Given their observations that constitutive PD-1 pathway deficiency impaired memory formation, Pauken, Godec, and Odorizzi et al. wondered whether the same effect might occur in temporary PD-1 axis blockade. The effects of antibody blockade of PD-1 in WT mice early during infection with X31-GP33 (day 1 through day 8 p.i.) were similar to those observed in PD-1 knockouts during the effector phase; however, the differences were not sustained at the memory time point. At day 60, while PD-L1/L2 DKO mice showed a quantitative defect in memory recall responses, WT mice treated early with anti-PD-1 showed normal memory responses, suggesting that short-term PD-1 blockade during priming did not induce the same long term defects in the generation of memory T cells.
Overall, Pauken, Godec, and Odorizzi et al. showed that PD-1 plays an important role in the development of both early effector T cell responses and long-term memory T cell responses following acute infection. While constitutive deficiency of the PD-1 axis led to early and enhanced effector responses during initial infection, it also increased cell death and diminished memory recall responses. Interestingly, the same long-term effects were not observed when PD-1 was temporarily blocked during the priming phase only, suggesting that the timing and/or the duration of the loss of PD-1 signaling were important, which could be relevant in the context of optimizing the scheduling of PD-1 blockade for the treatment of cancer.
by Lauren Hitchings
Meet the researcher
This week, first co-author Kristen E. Pauken and lead author Arlene H. Sharpe answered our questions.
What prompted you to tackle this research question?
A.H.S./K.E.P.: The PD-1 pathway has a fundamental role in mediating T cell exhaustion, but how this pathway regulates acute infection and memory development is poorly understood. PD-1 pathway blockade is FDA-approved for treating over 20 types of cancer, so understanding how the PD-1 pathway controls effector and memory T cell differentiation has therapeutic significance. Since PD-1 is expressed by all T cells during acute activation, and the cues CD8+ T cells receive during activation are critical for shaping both effector and memory T cell responses, we hypothesized that the PD-1 pathway would play a role here. We found that permanent loss of PD-1 signals led to improved effector responses, but impaired memory responses during acute influenza infection.
What was the most surprising finding of this study for you?
A.H.S./K.E.P: Historically, loss of PD-1 signals has been associated with substantial improvements in CD8+ T cell responses in the context of chronic infection. Consequently, we were incredibly surprised when we observed the dramatic memory defect in the complete absence of PD-1 signals during acute influenza. Our findings have important implications for vaccines and for combining PD-1 blockade with other therapies, and we have further studies in progress to understand how the timing of PD-1 pathway blockade influences the generation and durability of memory responses.
What was the coolest thing you’ve learned (about) recently outside of work?
K.E.P.: As many during the COVID-19 pandemic, my spouse and I have been faced with an unexpected increase in quality time with our toddler. This came two weeks after giving birth to our second child, so our transition to a family of four was not nearly as calm as expected. After many lessons in juggling, I have learned so much about the strength, resilience, and joy of my kids and spouse. It has been particularly amazing to watch the dynamic emotional range of my toddler, the huge ups and downs. I’ve learned to embrace the chaos, especially when frosting and sprinkles are involved. It reminds me why I continue to pursue my passion of science, and to take a deep breath and smell the roses every once in a while.



